Mechanical cues from the extracellular matrix (ECM) microenvironment are known to be significant in modulating the fate of stem cells to guide developmental processes and maintain bodily homeostasis. Tissue engineering has provided a promising approach to the repair or regeneration of damaged tissues. Scaffolds are fundamental in cell-based regenerative therapies. Developing artificial ECM that mimics the mechanical properties of native ECM would greatly help to guide cell functions and thus promote tissue regeneration. In this review, we introduce various mechanical cues provided by the ECM including elasticity, viscoelasticity, topography, and external stimuli, and their effects on cell behaviours. Meanwhile, we discuss the underlying principles and strategies to develop natural or synthetic biomaterials with different mechanical properties for cellular modulation, and explore the mechanism by which the mechanical cues from biomaterials regulate cell function toward tissue regeneration. We also discuss the challenges in multimodal mechanical modulation of cell behaviours and the interplay between mechanical cues and other microenvironmental factors.
As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration.
In this review, we summarise recent findings in the area of cellular regulation and how it is affected by mechanical cues from biomaterials, including elasticity, viscoelasticity, topography, and external stimuli, and discuss how these mechanical cues are controlled and manipulated to promote tissue regeneration (Figure 1). We also discuss the interwoven effects, or multimodal mechanical regulation, on cell behaviours, and the interplay between matrix mechanical cues and other environmental factors.
Figure 1.
Schematic illustration of matrix mechanical cues that regulate cell behaviours. In general, the matrix mechanical cues include elasticity, viscoelasticity, topography, fibre stiffness, and external stimuli, all of which can regulate many cellular behaviours, including cell adhesion, spreading, proliferation, migration and differentiation. However, a single mechanical stimulus is generally insufficient to induce stem cell differentiation and achieve tissue regeneration; instead, multimodal mechanical factors including elasticity, viscoelasticity, topography and external mechanical stimuli would have a synergistic effect in guiding cell behaviours to promote tissue regeneration.
Cellular Modulation by Matrix Elasticity
Elasticity of natural tissues
Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration.
Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3).
Figure 3.
Stages in the development of research showing how matrix elasticity regulates cell behaviours. (A) The pioneering study demonstrates that human BMSCs were effectively induced to differentiate into neuronal, muscle or bone lineages when they were cultured on soft, medium, or stiff substrates. Stiffness was then considered as one of the most important mechanical cues in tissue engineering. Reprint from Engler et al.20 Copyright 2006, with permission from Elsevier. (B) In the following years, numerous biomaterials have been developed to explore cell behaviours including stem cell fate affected by substrate elasticity, and its underlying molecular mechanism. (C) In recent years, many studies began to focus on the interplay between substrate elasticity and other cues (such as topography, geometry, growth factors, etc.) and its effect on cell behaviours. In addition, to simulate dynamic changes in stiffness in vivo, biomaterials with dynamic elasticity in situ have been designed to explore mechanobiological pathways that may differ from those under static cell culture. Further, new mechanosensitive proteins have been found to be involved in the cellular responses toward matrix elasticity, including YAP, piezo, caveolin-1, etc. BMSCs: bone marrow mesenchymal stem cells; FAK: focal adhesion kinase; MAPK: mitogen-activated protein kinase; ROCK: RHO-related protein kinase 1; YAP: yes-associated protein.
In 2016, we summarized the general principles and strategies to develop natural or synthetic materials with different elasticities for cellular modulation.18 In this article, we discuss the underlying principles and recent advances in further understanding how biomaterials can be designed to control the elastic microenvironment and thus regulate cell behaviours toward tissue regeneration.
Biomaterials with adjustable elasticity
Numerous biomaterials with different elasticities have been fabricated to study the effects of ECM stiffness on various cell behaviours. The latest strategies to manipulate the elasticity of matrix for studying the effect of tissue elastic environment on cell behaviours have been summarized (Additional Table 1).
Many natural or synthetic biomaterials with adjustable elasticity have been developed for cell modulation.20-33 In order to expand the library of controllable elastic biomaterials, study the interplaying effect of matrix elasticity with other physical cues, and develop elastic materials with tissue heterogeneity, some new strategies have been developed recently. For example, to understand the cross-talk effect of porosity and elasticity on encapsulated mesenchymal stem cells (MSCs), Ansari et al.34 used microfluidic technology to prepare alginate microspheres with different elasticities and architectures which they used as the carrier of MSCs. The elasticity of alginate hydrogels increased with increased calcium ion concentration, while the porosity decreased. MSCs can be encapsulated into these microspheres uniformly, and be regulated by the interplaying effect of elasticity and microstructure. Enzyme-catalysed crosslinking of silk fibroin using horseradish peroxidase and hydrogen peroxide has been reported to form hydrogels with adjustable stiffness in the range of 0.2–10 kPa. Wang et al.35 recently introduced inert silk fibroin nanofibres into these gels, and significantly improved the elasticity up to 9–60 kPa to match the mechanical properties of many tissues, especially bone and cartilage. To mimic the gradual change in elasticity of the annulus fibrosus (AF) along the radial direction, we synthesized biodegradable poly(ether carbonate urethane)urea with diverse Young’s moduli (2.5–13.4 MPa), by adjusting the molecular weight of polydiols and the feed ratios of hard molecular segment to soft molecular segment.36
In natural tissues, ECM remodelling often changes the physical stiffness of the local environment. In the past, most research in this field was carried out using static substrates as artificial ECM. In recent years, some hydrogels with dynamic stiffness in situ have emerged to simulate the dynamic stiffness changes which occur in vivo. For example, Günay et al.37 used light-mediated anthracene group [4+4] photodimerization to prepare polyethylene glycol (PEG)-based hydrogels, which were cytocompatible and provided a tailorable initial modulus that could be further strengthened. To correspond to the stiffness of a healthy or a fibrotic heart, the hydrogels were stiffened from 10 kPa to 50 kPa to monitor the stiffness-dependent localization of nuclear factor of activated T cells. This type of material provides an intriguing way to reveal the dynamics of mechano-reactive signalling pathways, which differ from those under static cell culture. Moreover, Young and Engler38 prepared hydrogels with time-dependent stiffness by polymerization of thiolated hyaluronic acid and polyethylene glycol diacrylate (PEGDA) to simulate the dynamic mechanics of heart development. The stiffness of the hydrogel increased from 1.9 ± 0.1 kPa to 8.2 ± 1.1 kPa within 69.6 hours after polymerization, and ester hydrolysis had no significant effect on hydrogel stiffening over 2 weeks. Compared with the static polyacrylamide hydrogel group, pre-cardiac cells cultured on dynamic hyaluronic acid hydrogels formed more mature myocardial fibres and the expression level of mature cardiac-specific markers was significantly increased. More interestingly, Fu et al.39 used a redox-responsive protein folding switch to fabricate a dynamic protein hydrogel with a Young’s modulus which could be reversibly regulated using redox conditions. Whether starting from the reduced state or from the oxidized state, the Young’s modulus of the hydrogels was reversible cyclically in response to changes in the redox condition. Importantly, such reversibility remained even after storage in phosphate-buffered saline or cell culture medium for 2 weeks. They further found that human lung fibroblasts dynamically changed their morphology with the cyclic variation of the hydrogel’s Young’s modulus between ~6 kPa and ~20 kPa, demonstrating the potential of such material in mechanobiology studies.
Effects of matrix elasticity on cell behaviours
Cell adhesion
Cell adhesion to the ECM is a complex process, involving cell attachment, cell spreading, actin cytoskeleton organization and then focal adhesion (FA) formation. Cell attachment can be completed in a short time to connect cells to the matrix. The cells then spread on the matrix and develop different morphologies. Following that, actin polymerises into microfilaments to form the actin cytoskeleton. Finally, the formation of FAs constitutes the connection between the matrix and the cellular cytoskeleton. The FAs, mainly composed of transmembrane adhesion receptors including integrins, vinculins and paxillins, can transfer biological and mechanical stimuli into cells and initiate molecular pathways which subsequently regulate gene expression.20 A number of studies have demonstrated that the assembly and turnover of FAs are affected by the elasticity of the substrate. In general, cells have more stable and matured FAs on stiffer substrates, and have reduced and diffused adhesion complexes on softer substrates. For example, osteocytes form more stable vinculin-containing adhesion complexes on a stiff substrate than those on a soft substrate.40 Myocardial cells also exhibit a stronger traction force on stiffer substrates.41 However, the relationship between cell adhesion strength and matrix elasticity is non-linear. It has been reported that when fibroblasts and endothelial cells (ECs) are cultured sparsely without cell-cell contacts, the spread area of cells abruptly changes at a stiffness range of around 3 kPa.42
Cell migration
Cell migration is an essential cell behaviour in diverse biological processes including development, angiogenesis, wound healing and inflammation. It is also highly sensitive to environmental stiffness. For example, the migration speed of HT-1080 fibrosarcoma cells doubled on a stiff gel (13 kPa) compared to those on a soft gel (0.34 kPa).43 When renal progenitor cells were seeded onto substrates with different elasticities, cell migration was limited on the substrates of 0.5 kPa and 2 kPa, but increased on the substrate of 4 kPa and remained stable on the stiff plates.44 Migration speed thus increases or decreases with the change of matrix elasticity, in other words, many cell types have an optimal stiffness, at which cell migration is maximal. For instance, smooth muscle cells show obvious biphasic dependence on matrix elasticity, and have a maximal speed at a particular stiffness.45 To predict the optimal environmental stiffness for different cells, Bangasserere et al.46 developed a cell migration simulator and successfully determined the optimal stiffness for embryonic chick forebrain neurons (~1 kPa) and U251 glioma cells (~100 kPa). The simulator would help researchers find the optimum stiffness to encourage cells to move to damaged or unhealthy tissues to restore the function of the tissue. Matrix elasticity affects not only the ability of cells to migrate, but also their direction of migration. Directional migration has been observed from softer to more stiff regions for many cell types.47-49 For example, vascular smooth muscle cells showed obvious directional migration on a mechanical gradient matrix coated with fibronectin, but not on a mechanical gradient matrix coated with laminin, indicating that the adhesion ligand might be an important element which affects cell migration in response to mechanical gradients.47
Cell proliferation and apoptosis
Cell proliferation and apoptosis is the basis of growth, development and regeneration of organisms. There are many important factors that affect cell proliferation and apoptosis, and matrix elasticity is among them. For example, proliferation of periodontal ligament stem cells on stiff substrates was faster than on soft substrates.50 The effect of matrix stiffness on cell proliferation is especially prominent in tumour cells. Increased proliferation of breast cell lines has been seen on stiff substrates.51 Many cells divide much more rapidly on a relatively stiffer substrate, while some cells behave differently. For instance, neural stem/progenitor cells proliferated well on soft substrates (< 10 kPa) and exhibited maximal proliferation on 3.5 kPa substrates.52 Robinson et al.53 developed a series of PEG-based hydrogels to investigate the effect of substrate stiffness on various vascular cells, including human umbilical vein ECs, human aortic adventitial fibroblasts, and human aortic vascular smooth muscle cells. Proliferation of human aortic adventitial fibroblasts increased with the increased hydrogel elasticity, yet proliferation of human umbilical vein ECs demonstrated an opposite trend with most rapid proliferation on the softest hydrogels. Interestingly, proliferation of human aortic vascular smooth muscle cells was hardly affected by matrix stiffness. Therefore, the proliferative response of cells to matrix elasticity showed cell-type specificity and nonlinearity. Accordingly, a range of substrate stiffness should be tested to determine the optimum moduli for cell growth. ECM with appropriate stiffness may promote cell proliferation and slow apoptosis.
Cell differentiation
Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58
Implications for tissue regeneration
Matrix elasticity has a great impact on cell fate and activity. Therefore, this feature must be considered carefully when developing biomaterials for tissue engineering. Choosing suitable matrix stiffness to promote tissue regeneration has been widely used in the regeneration of tissues including nerve, muscle, bone, cartilage, and blood vessels. For example, the formation and activity of neuronal networks can be modulated by matrix stiffness.59 On soft substrates, the migration of cortical neurons is enhanced, which leads to a faster formation of neuronal networks, while on stiff substrates, the pre-synaptic density, number of action potentials and miniature synaptic currents are enhanced, which indicates that matrix stiffness should be a key design parameter in scaffold fabrication for neural tissue engineering. However, for bone tissue engineering, a stiffer substrate is more appropriate. For example, three-dimensional (3D) porous collagen/hydroxyapatite scaffolds with the same microstructures but different stiffnesses were fabricated for bone tissue engineering.60 Bone matrix proteins, including osteocalcin and osteopontin, exhibited significantly higher expression on the highest stiffness group after 3 weeks of culture. The expression of osteocalcin and osteopontin was also higher on the highest stiffness group after 1 and 3 months of implantation in vivo. Matrix elasticity also plays a critical role in regulating chondrocyte behaviour and phenotype in vitro for cartilage tissue engineering. Many key features of chondrocytes including cell spreading area, percentage of irregular cell shape distribution, and mechanical characteristics are matrix stiffness-dependent,61 implying that choosing the optimal matrix elasticity is very important for cartilage tissue engineering.62-64 For instance, architectured 3D hydrogel scaffolds (ArcGels) were used to explore the stiffness effect on human articular chondrocytes in vitro and in vivo. The results showed that chondrogenic differentiation was more efficiently promoted on soft ArcGel in vitro, while the chondrogenic phenotype of human articular chondrocytes and retention of cartilaginous matrix were better maintained on stiff ArcGel upon ectopic implantation. In general, when the stiffness of a tissue engineering scaffold is similar to that of the host tissue, the stem cells tend to proliferate, secrete a large amount of ECM and differentiate into the cells of the specific tissue.
Cellular Modulation by Matrix Viscoelasticity Viscoelastic property of living tissues
Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation.
Biomaterials with adjustable viscoelasticity
Cells reside in a highly-dynamic, viscoelastic ECM, read microenvironmental cues and respond to them in a mechanically-sensitive manner to maintain the health of cells and tissues. In order to study the correlation between cell function and viscoelasticity, many hydrogels with adjustable viscoelasticity have been developed to simulate the microenvironment by matching the natural viscoelastic ECM. There are various molecular origins of viscoelastic hydrogels, including the movements of entangled polymers or “loose ends” of crosslinked polymers, protein folding, and deriving from physically non-covalent crosslinkers or chemically-dynamic covalent crosslinkers (Figure 4).
Figure 4.
Schematic diagram showing the molecular mechanism of the changes undergone by viscoelastic hydrogels when subjected to an external force. (A) Polyacrylamide-based hydrogels with different loss moduli varied through the movement of loose ends of polymer chains, or the loosing of entangled linear polyacrylamide. (B) Physically cross-linked hydrogels with varying viscoelasticity through the breaking of ionic interactions, hydrogen bonding, guest-host interactions, etc. In particular, for ionically cross-linked hydrogels, the viscoelasticity can also be tuned by incorporating covalent cross-linkers and polymer spacers. (C) Chemically-dynamic cross-linked hydrogels which change through the dissociation of chemical covalent bonds.
Polyacrylamide hydrogels, with varying loss moduli caused by movement of the loose ends of polymer chains, was an early approach to explore the effect of viscoelasticity on hMSC behaviour. The results highlighted that substrate creep was an important mechanical property of a biomaterial system to encourage cell growth and differentiation.70 Recently, Charrier et al.71 reported a type of soft, viscoelastic polyacrylamide hydrogel with independently-tuneable elasticity and viscous dissipation, due to the movements of entangled linear polyacrylamide which was sterically entrapped in the polyacrylamide crosslinked networks. They demonstrated that viscous dissipation is a determinant of cell phenotype and tissue homeostasis.71
However, most viscoelastic adjustable hydrogels were developed by creating networks with specific crosslinkers including physically non-covalent crosslinkers (e.g., ionic interaction,72-75 hydrogen bonding,76 guest-host interactions,77 etc.) and chemically dynamic covalent crosslinkers (e.g., acetyl exchange, disulphide bonds, hydrazone/oxime exchange, Diels-Alder reaction, etc.)75, 78-85 (Additional Table 2).
Alginate, a natural material, has been used to fabricate viscoelastic matrices for cell culture. The viscoelasticity can be controlled by changing the ratio of ionic cross-linker to covalent cross-linker, whose stress relaxation occurrs as a result of crosslinker unbinding.86 Based on this, Chaudhuri’s group.73, 87 reported that alginate hydrogels with various stress relaxation properties were obtained by changing the molecular weight of alginate or covalent incorporation of short PEG chains, which modulated the density of ionic crosslinks or led to a looser network in gels. Most recently, the same group found that stress relaxation of alginate hydrogel was simply and precisely modulated by changing the molecular weight and concentration of PEG.74 Increasing PEG concentration and molecular weight led to faster stress relaxation, higher loss modulus and increased creep. In order to simulate the fibrillar architecture of natural ECM, an interpenetrating network of click-functionalised alginate and fibrillar collagen type I was fabricated as a viscoelastic artificial ECM.75 Timing sequential ionic and covalent crosslinking allowed collagen type I to self-assemble into fibrous structures within the network. The stiffness and viscoelasticity was tuned by changing the mode and magnitude of crosslinking, without changing the micro-scale architecture of the hydrogels. The above viscoelastic alginate hydrogels were mostly ionically cross-linked, mainly using physical approaches to regulate viscoelasticity. Besides the ionic interaction, other physically cross-linked hydrogels have also been explored to form viscoelastic hydrogels including hydrogen bonding and guest-host interaction.
In addition, dynamically-reversible cross-links in hydrogels can undergo rapid breakage and re-formation when subjected to stress. The viscoelasticity of hydrogels can be adjusted by varying the density of dynamic chemical cross-linkers or adjusting the exchange rate of reversible cross-linkers. Imine and hydrazone bonds are reversible bonds which have been successfully used to fabricate many viscoelastic hydrogels, including hyaluronic acid, PEG, and alginate. For example, McKinnon et al.83 developed PEG-based viscoelastic hydrogels based on reversible hydrazone bonds with two different affinities. The stress relaxation rate (τ1/2) was tuned in the range of ~5–33,600 seconds, by changing the arm number of PEG or the ratio of aliphatic aldehyde to benzaldehyde crosslinkers. Further, the same group prepared 8-arm PEG viscoelastic hydrogels with stress relaxation times in the range of three orders of magnitude to check the effect on chondrocyte functions for cartilage tissue engineering. Recently, Sánchez-Morán et al.82 fabricated novel alginate hydrogels with tuneable stress relaxation based on reversible oxime bonds. They first prepared alkoxy-amine functionalized alginate, and then mixed this with aldehyde-containing alginate to form oxime cross-linked hydrogels. The stress relaxation and mechanical properties were adjusted by changing the concentration, mixing ratios, degree of oxidation of aldehyde-containing oxidized alginate or environmental factors (e.g. pH, temperature, and use of catalyst). Most interestingly, this study provided novel methods of preparing viscoelastic hydrogels in microbeads or microthread geometries.82 There are also other reversible covalent crosslinking methods, e.g. thioester, boronate ester, or Diels-Alder reaction, which can be used to fabricate dynamic hydrogels. For example, Brown et al.84 presented a viscoelastic hydrogel based on thioester exchange. The τ1/2 was tuneable across several orders of magnitude by controlling gel stoichiometry, pH, and crosslinker structure.
Although many strategies have been developed to tune the viscoelasticity of hydrogels as mentioned above, there is still a huge demand to create biomaterials with tuneable viscoelasticity to mimic the features of native tissues. Recently, in order to simulate the dynamic and heterogenous nature of tissues, Hui et al.80 developed a spatiotemporal controllable viscoelastic hydrogel based on phototunable hyaluronic acid, which showed great promise in simulating the range of viscoelasticities representing healthy and diseased cellular environments.
Effects of matrix viscoelasticity on cell behaviours
Cell adhesion
Recent studies showed that cell adhesion can be influenced by the viscoelasticity of the substrate as well its elasticity. For example, when U2OS cells were cultured on stress-relaxed alginate hydrogels, greater cellular FA formation was observed compared with that on purely elastic substrates, as indicated by the punctate localization of paxillin.72 Subsequently, the same group studied cell adhesion of MSCs in RGD-coupled alginate hydrogels with various rates of stress relaxation. They found that localization of integrin-β1 was increased at the periphery of the cell in faster relaxation gels.73 However, at any level of stress relaxation, there was no localisation of paxillin to the periphery of the cells, indicating that no conventional FAs were formed in this 3D viscoelastic hydrogel. In another study, the same group developed a novel viscoelastic matrix composed of hyaluronic acid and collagen type I,81 and found that FAs were promoted with higher expression of integrin and paxillin, while they were minimally-expressed and diffuse in the corresponding elastic substrate, due to the synergistic effect of viscoelasticity and fibrillar architecture. However, there have been some contradictory findings in other viscoelastic systems. In 2019, an alginate-PEG hydrogel with tuneable stress relaxation was developed for 3D culture of MSCs.74 It was found that localisation of integrin-β1 increased with faster relaxation, exhibiting higher levels of phosphorylated FA kinase (FAK) and paxillin. On polyacrylamide-based hydrogels with similar initial storage moduli and various loss moduli, paxillin clusters were much smaller on viscoelastic gels than on elastic gels, suggesting that energy dissipation of the substrate inhibits formation of FAs.70
Cell spreading
When a cell attaches to a substrate, it receives signals via integrins, after which actin polymerises to cause movement and extension of the cell membrane, while myosin contraction and membrane tension forces the cell to maintain tensional homeostasis, determining the cell spread area. A number of efforts have been devoted to investigating the effect of viscoelasticity on cell spreading in both two-dimensional (2D) and 3D culture. On 2D viscoelastic substrates, cell spreading was increased compared with elastic substrates, e.g. MSCs on collagen type I-coated polyacrylamide hydrogels, or U2OS cells72 and myoblasts83 on Arg-Gly-Asp (RGD)-modified alginate hydrogels. Conversely, when NIH-3T3 fibroblasts were cultured on boronate ester-based viscoelastic hydrogels, in all of the cell area and nuclear area, FA was lower than that on elastic hydrogels with the same storage modulus. In addition, with the change of stress relaxation time, the morphology of the cells will also change. Mostly, cells spread better and develop a less-rounded shaped on a fast stress relaxing substrate than on a slow stress relaxed substrate. For example, on an RGD-modified viscoelastic alginate hydrogel, fibroblasts and BMSCs showed greater cell expansion when τ1/2 was reduced from 1 hour to 1 minute.73 In 3D viscoelastic substrates, a similar phenomenon was observed. For example, in an RGD-coupled alginate-PEG hydrogel, the spreading of 3T3 fibroblasts was inhibited and cells with a spherical morphology were observed in slow stress relaxed gels, while cells spread more in the fast stress relaxing gels.73
Cell proliferation
The proliferation of various resident cells in a viscoelastic microenvironment was also found to be enhanced and regulated. Several studies demonstrated that hydrogel viscoelasticity has an obvious effect in promoting spreading and proliferation of fibroblasts in 2D or 3D culture.73, 88, 89 Similarly, when hMSCs were cultured on polyacrylamide-based viscoelastic hydrogels, the proliferation rate of hMSCs was significantly promoted by faster stress relaxation times.64 On an oxime cross-linked alginate hydrogel,82 2PK3 cells showed the same trend, with cells growing better in hydrogels with fast stress relaxation. All results indicate that stress relaxation time-dependent regulation may have beneficial effects on proliferation.
Cell migration
There is evidence that substrate stiffness guides cell migration, as does substrate viscoelasticity. Some studies have shown that mechanical cues from a viscoelastic matrix provide the orientation for migration. For instance, epithelial monolayer cells showed a special coalescence on a viscoelastic substrate, which was not observed on elastic materials.90 Sarcoma cells showed a faster migration speed in a collagen-based hypoxic hydrogel with faster stress relaxation than that in a gel with lower stress relaxation.89 Although it has been reported that cell migration can be regulated by the viscoelasticity of 2D or 3D substrates, the mechanism of this process needs to be further elucidated.
Cell differentiation
Stem cells have similar reactions to viscoelasticity in terms of cell spreading, proliferation, and migration. However, stem cells are unique in development and differentiation. Viscoelasticity has gained researchers’ attention recently for its role in directing stem-cell fate,14, 91 as well as elasticity emphasized in cell differentiation.
When hMSCs were cultured on a polyacrylamide gel system with varying loss modulus, the multilineage differentiation of hMSCs was enhanced in specific inductive media on high-loss modulus substrates. These results highlighted that substrate loss modulus was a significant mechanical parameter to be tailored for biomaterials in tissue engineering.70 Ionically-crosslinked alginate hydrogels are a typical viscoelastic hydrogel. A previous study found that MSCs undergo major adipogenic differentiation when the initial modulus is 1–10 kPa, and osteogenic differentiation when the initial modulus is 11–30 kPa. Chaudhuri et al.73 developed viscoelastic alginate hydrogels with various stress relaxation properties and initial elastic moduli, to study the differentiation of encapsulated MSCs. They found that MSCs exhibited primarily adipogenic differentiation at all timescales of stress relaxation when the initial elastic modulus was ~9 kPa, while the level of adipogenesis was decreased in rapidly-relaxing gels. In contrast, no adipogenic differentiation was observed when the initial elastic modulus was ~17 kPa, and osteogenic differentiation was significantly enhanced in faster stress relaxing gels. In addition, an interconnected bone-like matrix was formed in rapidly-relaxing gels with relaxation time at ~1 minute, indicating that faster stress relaxation promoted both osteogenesis and bone-forming activity. A subsequent study confirmed that faster relaxation also led to enhanced osteogenic differentiation of MSCs on PEG covalently-grafted alginate-PEG hydrogels.74
In summary, viscoelasticity of substrates has proven to be an important mechanical parameter in the regulation of stem cell behaviour. However, the library of viscoelastic hydrogels and the types of stem cells in the study were limited. Much more research should be undertaken to thoroughly understand the underlying mechanisms and rules of viscoelasticity on stem cell behaviour to facilitate the design of biomaterials for better tissue regeneration.
Implications in tissue regeneration
The viscoelastic properties of the ECM have been elucidated as a regulator of cell behaviours such as FA formation, cell spreading, proliferation, migration, and differentiation. Recently, there are some studies which showed the biological impacts of viscoelasticity in vivo. For example, Ghosh et al.92 prepared an ionically-crosslinked chitosan hydrogel with viscoelasticity to repair osteochondral defects in rabbits. They found that strain-reversible gels supported early cartilage-specific matrix formation and woven bone deposition. In a following study, the same group introduced stable covalent bonds into the gels to improve mechanical stability in vivo, and found that dual cross-linked chitosan hydrogels exhibited more collagen and mineral deposition compared to single ionically-crosslinked gels.93 The τ1/2 of a substrate has been proven to regulate many cell behaviours including stem cell differentiation in vitro. Mooney’s group addressed the hypothesis that substrate stress relaxation time could affect bone regeneration in vivo. They prepared alginate gels with various stress-relaxation times, which they then implanted into rat calvarial defects together with hMSCs.94 After 3 months of implantation, the rats that received fast-relaxing hydrogels (τ1/2 ≈ 50 seconds) showed much more growth of new bone compared with the rats that received slow-relaxing, stiffness-matched hydrogels. Strikingly, fast-relaxing hydrogel without encapsulated MSCs also promoted new bone formation, which was only slightly less than that induced by fast-relaxing hydrogels with MSCs (Figure 5). These results suggested that the viscoelasticity of biomaterials was an effective regulator of tissue regeneration in vivo, just like chemical and biological cues. Tissue engineering provides a promising solution for repair or regeneration of cartilage defects. To improve regenerative outcomes, matrices have been designed to incorporate viscoelastic features, making them much more similar to the mechanical features of native cartilage. To give one example, Lee et al.87 tested the impact of hydrogel stress relaxation rates on chondrocytes in 3D culture for cartilage tissue engineering. They found that faster relaxation promoted a significant increase in volume of interconnected cartilage matrix, while slower relaxation resulted in increased interleukin-1β secretion. In a recent study, Richardson et al.95 developed hydrazone crosslinked PEG hydrogels with tuneable viscoelastic properties for cartilage tissue engineering. They found that after 4 weeks of encapsulation, chondrocytes had more interconnected articular cartilage-specific matrix in stress-relaxation hydrogels. Collagen and sulphated glycosaminoglycans were deposited most in the adaptable hydrogel (22% benzyl-hydrazone hydrogel). Most recently, Li et al.96 used double network (DN) hydrogels based on gellan gum and polyethylene glycol acrylate (GG/PEGDA/DN) with stress-relaxing behaviour to study the effect on chondrogenic differentiation of BMSCs in a subcutaneous implant model. The results indicated that GG/PEGDA/DN provided a good physical environment for cell proliferation and spreading, and induced BMSCs to differentiate into the chondrogenic lineage, as demonstrated by a much greater level of collagen type II and transforming growth factor-β1. All these results show that stress relaxation of a substrate can mediate scaffold remodelling and tissue formation, which provides a new parameter to optimize tissue regeneration.
Figure 5.
Substrate stress-relaxation regulates scaffold remodelling and bone formation in vivo. (A) Young’s modulus and stress relaxation of slow- and fast-relaxing alginate hydrogels. (B) Representative micro-computed tomography renderings of rat calvaria 3 months post-injury. (C) Masson’s trichrome staining of the defect site in fast-relaxing and slow-relaxing gel conditions. Scale bars: 1 cm in B and 2 mm in C. Data are expressed as mean ± SD (n = 8–10) and were analysed by Student’s t-test. τ1/2: stress relaxation rate. Reproduced with the permission of Darnell et al.94 Copyright Wiley-VCH Verlag GmbH & Co. KGaA.
Comparison of Cellular Modulation Mediated by Elasticity and Viscoelasticity
In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours.
Cellular Modulation by Other Mechanical Cues
Effects of matrix topography on cell behaviours
A growing number of studies have shown that cell fate can be directed by the stiffness or viscoelasticity of the ECM. However, the ECM of various natural tissues such as bone, nerves, skin, kidney, or AF, has distinct topography at nano- or micro-scales.10, 102-105 Topographic factors including size, shape, and geometric arrangement, also considered to be an important biophysical cue, have substantial influence on cell behaviours.
Among these factors, size appears to play a critical role in modulating cell behaviours. For example, Nguyen et al.105 provided a comprehensive survey on the significance of its effect on cell adhesion, cell morphology and alignment, and neurite guidance. At both the nano- and micro-scale, cell behaviours can be significantly influenced by the size of topographical features. At the same time, the response of cells is diverse and strongly dependent on the type of cells. Cell behaviours often occur at both the nano- and micro-scales in biological interfaces and natural tissues, so an ideal artificial ECM should be biomimetic to match both size-scales and thus achieve a superior effect. For instance, ECs exhibited enhanced elongation when cultured on oriented nanofibrous scaffolds with different micro-grooved topographies.103 Recently, a 3D biomimetic nano-micro fibrous woven fabric/hydrogel composite scaffold was developed for heart valve engineering.106 The developers found that the native-like anisotropic features offered appropriate microenvironmental cues for cell proliferation and ECM deposition, representing a promising strategy to treat heart valve injury.
Alignment, one of the geometric arrangements, also exerts a great effect on cell behaviours. For cell adhesion, it was found that FAK phosphorylation was significantly increased in MSCs on aligned nanofibres compared with random nanofibres.105 For cell spreading, it was found that the cytoskeleton of human umbilical vein ECs on aligned fibres was stretched along the longitudinal direction of the fibre, while it extended in all directions on non-aligned fibres.107 Interestingly, cell alignment showed a more significant effect on micro-aligned scaffolds than on nano-aligned scaffolds.106 For cell migration, compared with random fibres, human adipose-derived stem cells cultured on aligned nanofibres exhibited a faster migration rate and higher synthetic ability of critical ECM molecules, such as collagen type I, tropoelastin, and matrix metalloproteinase-1.108 The effect of the alignment of electrospun polycaprolactone-gelatine ultrafine fibres on cell migration was also studied, and showed that the cell migration rate was faster in the direction of aligned fibres compared with non-aligned fibres.109 In addition, alignment affected cell proliferation and differentiation and was cell-type dependent. For example, aligned scaffold enhanced cell proliferation of keratocytes, while randomly-oriented scaffold enhanced the proliferation of corneal epithelial cells.110 The osteogenic differentiation of MSCs could also be modulated by fibre alignment. Alkaline phosphatase and calcium nodules were evenly distributed along the direction of aligned fibres, while gene expression of osteocalcin and osteopontin was significantly enhanced, indicating that aligned fibres were superior to non-aligned fibres in inducing osteogenic differentiation.107
Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces.
In summary, topography of a substrate, as a physical cue, shows great ability to modulate cell behaviour, and further, the combination of different topographical factors or other physical cues such as stiffness can achieve synergetic effects on cell fate. These findings provide guidance for the design of artificial ECM for use in tissue engineering and regenerative medicine.
Effects of fibre stiffness on cell behaviours
In native tissues, structural ECM proteins such as fibrillar collagens always range in diameter from 50 to 500 nm. To mimic the ECM structures, fibres have become the most commonly used scaffolds in tissue engineering. Fibres not only provide biological cues for embedded cells, but also serve as a source of nonlinear elasticity within the scaffold, with a unique ability to affect cell-cell communication and arrangement, as well as the degree of cell contraction. As mentioned above, substrate stiffness shows a great effect on various cell behaviours, so it is also very important to test the effect of the stiffness of fibres on cell behaviours. Recently, many studies have shown that the stiffness of fibrous scaffolds can affect cell morphology and signalling, and thus affect cell behaviour, including cell proliferation, adhesion, and migration.
Aligned ultrafine fibres have shown great potential in the construction of vascular grafts, simulating the anisotropic features of natural blood vessels. To investigate how the stiffness of aligned fibres influences the functionality of vascular cells, Yi et al.29 generated highly-aligned ultrafine fibres of poly(L-lactideco-caprolactone)/poly(L-lactic acid) with a stiffness range from 0.09 to 13.18 N/mm. They found that increased stiffness did not exert a significant effect on cell morphology or orientation, but inhibited cell adhesion, promoted cell migration and proliferation, and enhanced the assembly of F-actin fibres in human umbilical artery smooth muscle cells. Moreover, higher fibre stiffness significantly down-regulated contraction markers, and up-regulated gene expression of osteopontin and inflammatory genes, which indicated that higher fibre stiffness led to proliferative and pathological states of human umbilical artery smooth muscle cells.
Numerous studies have indicated that increased surface nano-roughness or stiffness of electrospun fibrous scaffolds is conducive for an osteogenic response. Recently, Jahanmard et al.114 incorporated multiwall carbon nanotubes into electrospun polycaprolactone nanofibres to adjust surface nanoroughness and stiffness independently. They found that osteoblast differentiation was improved by both these crucial parameters, yet through different mechanisms.114 All of these studies help us to understand the mechanical and biological responses of bone progenitor cells, and ultimately guide us to design better scaffolds for bone tissue engineering.
Fibrous scaffolds are also commonly being explored to restore the function and organization of dense connective tissues such as tendon, knee meniscus, and cartilage.115, 116 However, the lower porosity of electrospun fibres results in poor cell invasion, hindering cell migration into scaffolds. To test the impact of stiffness of electrospun fibres on cell invasion into a fibrous scaffold, Song et al.117 fabricated stiff and soft electrospun fibrous networks using crosslinked methacrylated hyaluronic acid. Enhanced meniscal fibrochondrocyte invasion and greater collagen deposition were observed on stiffer fibrous networks when compared with softer fibrous networks because of their easy deformation and increase in density under the influence of cell traction. All these results demonstrated that the stiffness of fibrous scaffolds is a key parameter in the design of scaffolds for tissue regeneration.
In addition, the stiffness of fibrous scaffolds also plays an important role in disease development processes or tissue repair processes. For example, our group recently reported that fibre stiffness of polyurethanes and fibre diameters showed a synergistic effect on the differentiation of AFSCs. AFSCs had a strong tendency to differentiate into various regional AF-like cells when cultured on a scaffold with physical cues similar to native AF tissue.102 Davidson et al.118 seeded hepatic stellate cells onto soft or stiff fibrous hyaluronic acid hydrogels, and found that cell spreading, α-smooth muscle actin expression, and formation of multicellular clusters decreased on stiff fibres compared with those on soft fibres, indicating the important influence of fibre stiffness on cell-material interactions and implying its relevance to disease and tissue repair processes.
External mechanical stimulation on cell behaviours
Accumulating evidence suggests that external mechanical stimulation regulates cellular processes and strongly influences cell fate.119 Stretching, compression, hydrostatic pressure and fluid shear force can affect cell self-renewal, differentiation, senescence and apoptosis, and play an important role in physiological and pathological processes.11 Indeed, cells in living tissues are constantly exposed to various chemical and physical signals, and then integrate these signals to guide cell differentiation specifics, support morphogenetic processes and define tissue architecture during embryonic development.120 Intracardiac fluid forces change cytoskeletal structure and gene expression profiles of cardiac ECs to promote the morphogenesis of cardiac tissues.121 Tensile forces, generated by the stretching of bronchial epithelium during intrauterine respiration, support the development of pulmonary smooth muscle.122 Long-bone morphogenesis is dependent on both local morphogen gradients and mechanical forces which guide cell assembly and promote growth to higher-order structures.123
In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130
In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation.
Molecular Basis
A comprehensive understanding of the cellular response to ECM properties is critical for developing suitable biomaterials to mimic natural tissues and increase seeded cell growth. How substrate mechanical signals are translated into intracellular bio-signals needs to be thoroughly elucidated. Cells perceive the mechanical environment through cell-cell adhesion and cell-ECM interactions, mechanosensitive ion channels and their primary cilia.14 Several mechanotransduction pathways have been proven to transduce mechanical cues into biochemical signals including FAK activated by integrins, Rho/RHO-related protein kinase 1 (ROCK) and mitogen-activated protein kinase (MAPK) as mechanoresponsive signalling, and downstream mechanotransduction signalling such as YAP/PDZ-binding motif (TAZ) (Figure 6), as summarized in Hynes’s review.139
Figure 6.
YAP/TAZ mediated pathway through mechano-transduction. A cell probes its ECM mechanical environment via membrane receptors (e.g. integrins) to transmit force which regulates the stability of focal adhesion complexes containing focal adhesion kinase (FAK). FAK phosphorylates and activates mechanoresponsive signalling elements, such as mitogen-activated protein kinase (MAPK) and transforming protein Ras homolog gene family member A (RHOA). Simultaneously, the intracellular force regulates the nuclear translocation of transcription regulator such as yes-associated protein (YAP)/transcriptional co-activator with PDZ-binding motif (TAZ).
The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity.
The methods used to explore the molecular mechanism of how cells sense ECM viscoelasticity are similar to those used in studying how cells sense ECM elasticity. RHO family GTPases are significant regulators of the actin cytoskeleton and of mechanotransduction. It has been found that RHO-mediated contractility and independent actomyosin contractility regulates osteogenic differentiation.68, 152 Increased Rac1 activity was also reported to enhance smooth muscle cell differentiation.153 YAP/TAZ has been considered as key mediators in cell responses to substrate stiffness, such as that in viscoelastic regulation of cell behaviours. For example, stress relaxation of a substrate has been found to lead to increased nuclear translocation of YAP, resulting in increased U2OS cell proliferation.74 Further, the cell spreading, proliferation, and osteogenic differentiation of MSCs was enhanced on gels with faster stress relaxation, which was proven to be mediated through integrin-based adhesion, actomyosin contractility, and nuclear localization of YAP.68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume.
Other mechanical cues, including matrix topography, fibre stiffness, and external mechanical stimulation, also affect the gene expression and fate of cells through the above signalling pathways and their interplays. For example, with the increase of FAK phosphorylation, the neuronal differentiation of hMSCs was promoted on equally-spaced nanogratings with widths of 250145 to 500 nm,154 suggesting that FAK phosphorylation is a signal transducer between integrins and the cytoskeleton, transmitting nanotopographical stimuli to the nucleus through intracellular contraction.145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158
Multimodal Mechanical Regulation of Cell Behaviours
The natural environment of cells is complex, and it is unrealistic to completely imitate the mechanical properties of a cell’s natural environment. Therefore, in past years, many studies have explored the effects of a single mechanical cue on cell behaviours. However, any given single mechanical regulation is not enough to induce stem cell differentiation and complete tissue regeneration. Researchers began to try combining multimodal mechanical cues to regulate cellular behaviours, including elasticity, viscoelasticity, topography and external stimuli.
The effects of topography and viscoelasticity on the fate of human adipose-derived stem cells were studied by combining nanostructures with viscoelastic elements. The results showed that in multi-layer modified nanostructures, the differentiation of human adipose-derived stem cells towards chondrogenesis and osteogenesis was better than that towards adipogenesis.159 By analysing the cell arrangement and neural differentiation of human umbilical cord mesenchymal stem cells and the growth of nerve processes in vitro and in vivo, the effects of elasticity and aligned topography as combined mechanical cues on cell behaviours were studied.160 After 2 weeks of culture in medium lacking a soluble neurogenic inducer, low elasticity and arranged topography of a layered fibrin hydrogel showed a synergistic effect in promoting the differentiation of human umbilical cord mesenchymal stem cells into nerve cells when compared with random fibrin hydrogel or a culture plate. In order to simulate the dynamic extracellular environment of bone marrow cells in vivo, a bone marrow stem cell substitute matrix based on a physically cross-linked hydrogel was designed. Elasticity was regulated dynamically by chemical stimulation. Under frequent mechanical stress, hMSCs on this hydrogel maintained the expression of Stro-1 (a potential marker of pluripotency of MSCs) for more than 20 days, regardless of the elasticity of the matrix.161 Stiffness and surface topography of the substrate are two key characteristics at the cell-substrate interface that affect cell behaviour as mentioned before.10 From all these studies, we can conclude that multi-mechanical factors play a synergistic role in guiding cell behaviour, or that some mechanical factors would play a leading role.
Interplay between Matrix Mechanical Cues and Other Environmental Factors
Mechanical cues such as matrix stiffness can promote cell arrangement, morphological change and phenotypic variation. The application of biochemical cues such as growth factors can regulate cells by activating certain signal transduction pathways and directing them to intracellular and/or extracellular remodelling. The application of mechanical and biochemical cues to control cell behaviour is common in tissue engineering and regenerative medicine. More and more studies have explored the comprehensive effects of biochemical cues and mechanical cues on cell behaviour.
For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167
Concluding Remarks and Future Perspectives
Over the past few years, great advances in mechanobiology have been achieved. Various mechanical cues from the ECM have been shown to profoundly influence cell behaviours, including proliferation, adhesion, migration, apoptosis, and differentiation. In this review, we introduced different mechanical cues from the ECM and their interplay effect on cell behaviours, including elasticity, viscoelasticity, topography, and external stimuli. However, the native environment of cells is more complicated. Single mechanical regulation is not enough to induce stem cell differentiation and achieve satisfactory tissue regeneration. In recent years, researchers have begun to combine different mechanical cues to regulate cell behaviours toward synergistic effects. However, most of them are just a combination of two factors, which is still far away from the realistic mechanical environment of cells. Therefore, how to mimic the complicated native mechanical environment experienced by cells remains challenging. Integrating multimodal mechanical cues from artificial ECM would help better understand the effects of a realistic mechanical environment on cell behaviours, which may lead to more effective tissue engineering. In addition, ECM remodelling in native tissues often changes the cells’ mechanical environment. There is growing interest in simulating the dynamic stiffness changes in vivo to study the mechanobiology. In recent years, some hydrogels with dynamic stiffness have been developed for regulating various cellular behaviours, yet the number of such hydrogels is still limited. In particular, hydrogels with directional and reversible changes in mechanical properties are very rare. Further expanding the application in a wide range of mechanobiology studies using these hydrogels was interesting and critical for the successful design of biomaterials for use in tissue regeneration. Viscoelasticity is another important mechanical property in living tissues and organs, which plays an important role in maintaining the physiological functions of organs, and especially in protecting tissues under persistent or irregular high stress. Several highlighted studies have found that hydrogels with stress relaxation or creep behaviours showed potent impacts on many cell behaviours. However, the role of time-dependent mechanics on cell biology should be further clearly studied and explored, which would advance our understanding of cell-matrix interactions to guide the successful design of biomaterials for tissue regeneration.
The design of biomaterials can benefit from our growing knowledge of tissue mechanobiology. Clearly understanding the interaction between cells with their complicated and dynamic mechanical environment will guide us to make biomimetic materials which can integrate and even induce tissue regeneration. Consequently, ideal biomaterials for tissue engineering should mimic the natural mechanical properties by multimodal regulation, to recapitulate the intrinsic diversity or dynamic nature of native tissue. Future studies in this field will eventually shorten the gap between basic research and clinical application in tissue regeneration.
Author contributions
supervision, and resources: BL and CZ; original draft preparation and visualisation: CZ; partial investigation and original draft: CZ, QW, SW, FH, HW, WZ, QY, CL, LD, JW, LY; manuscript review and editing: CZ and QW. All authors approved the final version of the manuscript.
Financial support
This work was supported by the National Natural Science Foundation of China (Nos. 81925027, 81871805, 31530024, 81672213), National Key R&D Program of China (No. 2016YFC1100203), Jiangsu Provincial Clinical Orthopaedic Centre, Key Laboratory of Stem Cells and Biomedical Materials of Jiangsu Province and Chinese Ministry of Science and Technology of China, and the Priority Academic Program Development (PAPD) of Jiangsu Higher Education Institutions of China.
Acknowledgement
None.
Conflicts of interest statement
The authors declare no competing financial interest.
Editor note: Bin Li is an Editorial Board member of Biomaterials Translational. He was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal’s standard procedures, with peer review handled independently of this Editorial Board member and his research group.
Open access statement
This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Additional files
Additional Table 1: Commonly-used biomaterials with various elasticities and their effects on cells.
Additional Table 2: Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Additional Table 1.
Additional Table 1.Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Additional Table 2.Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Targeting downstream subcellular YAP activity as a function of matrix stiffness with Verteporfin-encapsulated chitosan microsphere attenuates osteoarthritis
Mi,H. Y.; Jing,X.; Yilmaz,G.; Hagerty,B. S.; Enriquez,E.; Turng,L. S.
In situ synthesis of polyurethane scaffolds with tunable properties by controlled crosslinking of tri-block copolymer and polycaprolactone triol for tissue regeneration
Biomaterial arrays with defined adhesion ligand densities and matrix stiffness identify distinct phenotypes for tumorigenic and nontumorigenic human mesenchymal cell types
Macrí-Pellizzeri,L.; Pelacho,B.; Sancho,A.; Iglesias-García,O.; Simón-Yarza,A. M.; Soriano-Navarro,M.; González-Granero,S.; García-Verdugo,J. M.; De-Juan-Pardo,E. M.; Prosper,F.
Substrate stiffness and composition specifically direct differentiation of induced pluripotent stem cells
Respective effects of gelatin-coated polydimethylsiloxane (PDMS) substrates on self-renewal and cardiac differentiation of induced pluripotent stem cells (iPSCs)
Chondrogenesis of human bone marrow mesenchymal stem cells in 3-dimensional, photocrosslinked hydrogel constructs: Effect of cell seeding density and material stiffness
The use of nanoscale topography to modulate the dynamics of adhesion formation in primary osteoblasts and ERK/MAPK signalling in STRO-1+ enriched skeletal stem cells
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
Local 3D matrix microenvironment regulates cell migration through spatiotemporal dynamics of contractility-dependent adhesions
1
2015
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
Stem cell migration and mechanotransduction on linear stiffness gradient hydrogels
0
2017
Effects of substrate stiffness and cell-cell contact on mesenchymal stem cell differentiation
0
2016
Active mechanics and dynamics of cell spreading on elastic substrates
0
2014
Extracellular matrix stiffness causes systematic variations in proliferation and chemosensitivity in myeloid leukemias
1
2016
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
Remodelling the extracellular matrix in development and disease
1
2014
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
Tissue stiffness dictates development, homeostasis, and disease progression
4
2015
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
... Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration. ...
Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3). ...
... Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation. ...
Viscoelastic hydrogels for 3D cell culture
2
2017
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
... Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation. ...
Biophysical regulation of cell behavior-cross talk between substrate stiffness and nanotopography
4
2017
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
... A growing number of studies have shown that cell fate can be directed by the stiffness or viscoelasticity of the ECM. However, the ECM of various natural tissues such as bone, nerves, skin, kidney, or AF, has distinct topography at nano- or micro-scales.10, 102-105 Topographic factors including size, shape, and geometric arrangement, also considered to be an important biophysical cue, have substantial influence on cell behaviours. ...
... Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
... The effects of topography and viscoelasticity on the fate of human adipose-derived stem cells were studied by combining nanostructures with viscoelastic elements. The results showed that in multi-layer modified nanostructures, the differentiation of human adipose-derived stem cells towards chondrogenesis and osteogenesis was better than that towards adipogenesis.159 By analysing the cell arrangement and neural differentiation of human umbilical cord mesenchymal stem cells and the growth of nerve processes in vitro and in vivo, the effects of elasticity and aligned topography as combined mechanical cues on cell behaviours were studied.160 After 2 weeks of culture in medium lacking a soluble neurogenic inducer, low elasticity and arranged topography of a layered fibrin hydrogel showed a synergistic effect in promoting the differentiation of human umbilical cord mesenchymal stem cells into nerve cells when compared with random fibrin hydrogel or a culture plate. In order to simulate the dynamic extracellular environment of bone marrow cells in vivo, a bone marrow stem cell substitute matrix based on a physically cross-linked hydrogel was designed. Elasticity was regulated dynamically by chemical stimulation. Under frequent mechanical stress, hMSCs on this hydrogel maintained the expression of Stro-1 (a potential marker of pluripotency of MSCs) for more than 20 days, regardless of the elasticity of the matrix.161 Stiffness and surface topography of the substrate are two key characteristics at the cell-substrate interface that affect cell behaviour as mentioned before.10 From all these studies, we can conclude that multi-mechanical factors play a synergistic role in guiding cell behaviour, or that some mechanical factors would play a leading role. ...
Mechanobiology of YAP and TAZ in physiology and disease
2
2017
... As is well known, movement of the human body is realized through precise coordination of different systems, including physical movements guaranteed by the skeletal system, blood circulation generated by the circulatory system, and gas exchange carried out by the respiratory system. The mechanical loads generated by these physical activities regulate tissue homeostasis through biomechanical changes. The human body also responds to mechanical signals from the environment, such as bone microarchitecture remodelling under local mechanical stimulation.1 These macroscopic phenomena in living tissue are the result of an accumulated series of cellular reactions, especially the interaction between cells and their corresponding extracellular matrix (ECM). This interaction has become the critical mechanical force not only in maintaining homeostasis of the body, but also in regulating stem cell behaviours and fate to direct developmental processes. ECM, a network composed of a series of functional proteins and polysaccharides, transmits external mechanical stimuli to cells and regulates cell spreading, proliferation, differentiation, senescence or even carcinogenesis.2-6 Meanwhile, the resident cells can remodel their cytoskeleton as well as the composition and structure of the ECM in response to the mechanical microenvironment, resulting in substantial differences in tissue development, physiological activity, disease progression and tissue reconstruction.7 Thus, different tissues form unique ECMs with distinct mechanical properties including elasticity8 and viscoelasticity,9 various nano- or micro-topographies,10 and different external mechanical environments.11 Tissue engineering provides a promising approach to produce new viable tissue for repairing/replacing damaged tissues and thus restoring their biological functions. Since scaffolds are a fundamental component of cell-based regenerative therapies, it is important to understand how they transmit mechanical stimulation to cells. Meanwhile, it is also critical to explore the mechanisms which guide cell function by controlling the mechanical cues of naturally- or synthetically-derived biomaterials to promote tissue regeneration. ...
... Accumulating evidence suggests that external mechanical stimulation regulates cellular processes and strongly influences cell fate.119 Stretching, compression, hydrostatic pressure and fluid shear force can affect cell self-renewal, differentiation, senescence and apoptosis, and play an important role in physiological and pathological processes.11 Indeed, cells in living tissues are constantly exposed to various chemical and physical signals, and then integrate these signals to guide cell differentiation specifics, support morphogenetic processes and define tissue architecture during embryonic development.120 Intracardiac fluid forces change cytoskeletal structure and gene expression profiles of cardiac ECs to promote the morphogenesis of cardiac tissues.121 Tensile forces, generated by the stretching of bronchial epithelium during intrauterine respiration, support the development of pulmonary smooth muscle.122 Long-bone morphogenesis is dependent on both local morphogen gradients and mechanical forces which guide cell assembly and promote growth to higher-order structures.123 ...
Effects of extracellular matrix viscoelasticity on cellular behaviour
1
2020
... Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration. ...
Soft biological materials and their impact on cell function
2
2007
... Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration. ...
Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3). ...
Mechanical forces direct stem cell behaviour in development and regeneration
2
2017
... Stem cells have similar reactions to viscoelasticity in terms of cell spreading, proliferation, and migration. However, stem cells are unique in development and differentiation. Viscoelasticity has gained researchers’ attention recently for its role in directing stem-cell fate,14, 91 as well as elasticity emphasized in cell differentiation. ...
... A comprehensive understanding of the cellular response to ECM properties is critical for developing suitable biomaterials to mimic natural tissues and increase seeded cell growth. How substrate mechanical signals are translated into intracellular bio-signals needs to be thoroughly elucidated. Cells perceive the mechanical environment through cell-cell adhesion and cell-ECM interactions, mechanosensitive ion channels and their primary cilia.14 Several mechanotransduction pathways have been proven to transduce mechanical cues into biochemical signals including FAK activated by integrins, Rho/RHO-related protein kinase 1 (ROCK) and mitogen-activated protein kinase (MAPK) as mechanoresponsive signalling, and downstream mechanotransduction signalling such as YAP/PDZ-binding motif (TAZ) (Figure 6), as summarized in Hynes’s review.139 ...
Towards intervertebral disc engineering: Bio-mimetics of form and function of the annulus fibrosus lamellae
2
2019
... Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration. ...
Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3). ...
Altered cartilage mechanics and histology in knee osteoarthritis: relation to clinical assessment (ICRS Grade)
1
2005
... Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration. ...
Targeting downstream subcellular YAP activity as a function of matrix stiffness with Verteporfin-encapsulated chitosan microsphere attenuates osteoarthritis
1
2020
... Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration. ...
Cellular modulation by the elasticity of biomaterials
5
2016
... Over the last two decades, the mechanical parameter of living tissue that has received the most attention is elasticity. There are different factors, including stiffness, elastic modulus, flexibility, and rigidity, which characterize the resistance of materials to deformation. In fact, nearly all natural tissues are viscoelastic.12 To avoid confusion, the elasticity of natural tissues described in this section refers to the stiffness of natural tissues. Each tissue or organ in our body, from soft fat to rigid bone, has characteristic elasticity to fulfil specific physiological needs8, 13-15 (Figure 2A). For example, the elasticity of bone is much higher than that of other tissues, mainly because of its function in supporting the body and protecting other tissues. Changes of tissue elasticity in pathological states are usually related to the progression of disease. For instance, reduced cartilage stiffness is correlated with osteoarthritis progression. The stiffness of cartilage is 0.50 ± 0.14, 0.37 ± 0.13, and 0.28 ± 0.12 MPa for samples with International Cartilage Repair Society grades of 1, 2, and 3, respectively.16 Recently, Zhang et al.17 have confirmed that the human cartilage matrix hardens during osteoarthritis progression. On the other hand, a mechanotransducer-targeted drug which mediates ECM stiffness to maintain chondrocyte phenotype is able to prevent cartilage degeneration. Therefore, maintaining the elasticity of natural tissues is very important. Understanding how tissue elasticity regulates the behaviours of cells, especially stem cells, is essential for designing biomaterials to guide cell functions and lead to tissue regeneration.
Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3). ...
... Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3). ...
... In 2016, we summarized the general principles and strategies to develop natural or synthetic materials with different elasticities for cellular modulation.18 In this article, we discuss the underlying principles and recent advances in further understanding how biomaterials can be designed to control the elastic microenvironment and thus regulate cell behaviours toward tissue regeneration. ...
... Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation. ...
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
Tissue elasticity and the ageing elastic fibre
2
2009
... Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3). ...
... Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation. ...
... Every tissue is a specific combination of ECM, local cells and growth factors. Among them, the ECM is the most important regulator of tissue elasticity, and plays a significant role in modulating the behaviours of the resident cells, including adhesion, spreading, proliferation, migration, differentiation, and apoptosis.18 Changes of ECM stiffness will lead to changes of the physical properties of the tissue and consequently to changes in cell behaviours, which, in turn, may drive abnormal cell proliferation and tumourigenesis.19 In addition, stiffness which matches the native ECM may induce stem cells to differentiate into residential tissue cell lines. For example, Engler et al.20 have demonstrated that human bone marrow mesenchymal stem cells (BMSCs) can be induced into neuronal, muscle or bone lineages when cultured on soft, medium, or stiff hydrogel matrix, respectively. In recent decades, the effects of ECM stiffness on cell activities have been thoroughly explored. Numerous studies have revealed that matrix stiffness has a substantial impact on stem cell differentiation (Figure 2B). The development of this field can be divided roughly into three stages (Figure 3). ...
... 20 Copyright 2006, with permission from Elsevier. (B) In the following years, numerous biomaterials have been developed to explore cell behaviours including stem cell fate affected by substrate elasticity, and its underlying molecular mechanism. (C) In recent years, many studies began to focus on the interplay between substrate elasticity and other cues (such as topography, geometry, growth factors, etc.) and its effect on cell behaviours. In addition, to simulate dynamic changes in stiffness in vivo, biomaterials with dynamic elasticity in situ have been designed to explore mechanobiological pathways that may differ from those under static cell culture. Further, new mechanosensitive proteins have been found to be involved in the cellular responses toward matrix elasticity, including YAP, piezo, caveolin-1, etc. BMSCs: bone marrow mesenchymal stem cells; FAK: focal adhesion kinase; MAPK: mitogen-activated protein kinase; ROCK: RHO-related protein kinase 1; YAP: yes-associated protein.
In 2016, we summarized the general principles and strategies to develop natural or synthetic materials with different elasticities for cellular modulation.18 In this article, we discuss the underlying principles and recent advances in further understanding how biomaterials can be designed to control the elastic microenvironment and thus regulate cell behaviours toward tissue regeneration. ...
... Many natural or synthetic biomaterials with adjustable elasticity have been developed for cell modulation.20-33 In order to expand the library of controllable elastic biomaterials, study the interplaying effect of matrix elasticity with other physical cues, and develop elastic materials with tissue heterogeneity, some new strategies have been developed recently. For example, to understand the cross-talk effect of porosity and elasticity on encapsulated mesenchymal stem cells (MSCs), Ansari et al.34 used microfluidic technology to prepare alginate microspheres with different elasticities and architectures which they used as the carrier of MSCs. The elasticity of alginate hydrogels increased with increased calcium ion concentration, while the porosity decreased. MSCs can be encapsulated into these microspheres uniformly, and be regulated by the interplaying effect of elasticity and microstructure. Enzyme-catalysed crosslinking of silk fibroin using horseradish peroxidase and hydrogen peroxide has been reported to form hydrogels with adjustable stiffness in the range of 0.2–10 kPa. Wang et al.35 recently introduced inert silk fibroin nanofibres into these gels, and significantly improved the elasticity up to 9–60 kPa to match the mechanical properties of many tissues, especially bone and cartilage. To mimic the gradual change in elasticity of the annulus fibrosus (AF) along the radial direction, we synthesized biodegradable poly(ether carbonate urethane)urea with diverse Young’s moduli (2.5–13.4 MPa), by adjusting the molecular weight of polydiols and the feed ratios of hard molecular segment to soft molecular segment.36 ...
... Cell adhesion to the ECM is a complex process, involving cell attachment, cell spreading, actin cytoskeleton organization and then focal adhesion (FA) formation. Cell attachment can be completed in a short time to connect cells to the matrix. The cells then spread on the matrix and develop different morphologies. Following that, actin polymerises into microfilaments to form the actin cytoskeleton. Finally, the formation of FAs constitutes the connection between the matrix and the cellular cytoskeleton. The FAs, mainly composed of transmembrane adhesion receptors including integrins, vinculins and paxillins, can transfer biological and mechanical stimuli into cells and initiate molecular pathways which subsequently regulate gene expression.20 A number of studies have demonstrated that the assembly and turnover of FAs are affected by the elasticity of the substrate. In general, cells have more stable and matured FAs on stiffer substrates, and have reduced and diffused adhesion complexes on softer substrates. For example, osteocytes form more stable vinculin-containing adhesion complexes on a stiff substrate than those on a soft substrate.40 Myocardial cells also exhibit a stronger traction force on stiffer substrates.41 However, the relationship between cell adhesion strength and matrix elasticity is non-linear. It has been reported that when fibroblasts and endothelial cells (ECs) are cultured sparsely without cell-cell contacts, the spread area of cells abruptly changes at a stiffness range of around 3 kPa.42 ...
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
Fabrication and characterization of chitosan-hyaluronic acid scaffolds with varying stiffness for glioblastoma cell culture
1
2018
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Tunable fibrin-alginate interpenetrating network hydrogels to support cell spreading and network formation
1
2020
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
The influence of matrix stiffness on the behavior of brain metastatic breast cancer cells in a biomimetic hyaluronic acid hydrogel platform
1
2018
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Tunable stiffness of graphene oxide/polyacrylamide composite scaffolds regulates cytoskeleton assembly
1
2018
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Tunable and reversible substrate stiffness reveals a dynamic mechanosensitivity of cardiomyocytes
1
2019
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
In situ synthesis of polyurethane scaffolds with tunable properties by controlled crosslinking of tri-block copolymer and polycaprolactone triol for tissue regeneration
1
2018
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Silk fibroin/collagen protein hybrid cell-encapsulating hydrogels with tunable gelation and improved physical and biological properties
1
2018
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Three-dimensional encapsulation of adult mouse cardiomyocytes in hydrogels with tunable stiffness
1
2020
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Stiffness of aligned fibers regulates the phenotypic expression of vascular smooth muscle cells
2
2019
... Aligned ultrafine fibres have shown great potential in the construction of vascular grafts, simulating the anisotropic features of natural blood vessels. To investigate how the stiffness of aligned fibres influences the functionality of vascular cells, Yi et al.29 generated highly-aligned ultrafine fibres of poly(L-lactideco-caprolactone)/poly(L-lactic acid) with a stiffness range from 0.09 to 13.18 N/mm. They found that increased stiffness did not exert a significant effect on cell morphology or orientation, but inhibited cell adhesion, promoted cell migration and proliferation, and enhanced the assembly of F-actin fibres in human umbilical artery smooth muscle cells. Moreover, higher fibre stiffness significantly down-regulated contraction markers, and up-regulated gene expression of osteopontin and inflammatory genes, which indicated that higher fibre stiffness led to proliferative and pathological states of human umbilical artery smooth muscle cells. ...
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
The influence of the stiffness of GelMA substrate on the outgrowth of PC12 cells
1
2019
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Combinational effect of matrix elasticity and alendronate density on differentiation of rat mesenchymal stem cells
1
2015
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Decoupling the effects of stiffness and fiber density on cellular behaviors via an interpenetrating network of gelatin-methacrylate and collagen
1
2017
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Regulation of the fate of dental-derived mesenchymal stem cells using engineered alginate-GelMA hydrogels
2
2017
... Many natural or synthetic biomaterials with adjustable elasticity have been developed for cell modulation.20-33 In order to expand the library of controllable elastic biomaterials, study the interplaying effect of matrix elasticity with other physical cues, and develop elastic materials with tissue heterogeneity, some new strategies have been developed recently. For example, to understand the cross-talk effect of porosity and elasticity on encapsulated mesenchymal stem cells (MSCs), Ansari et al.34 used microfluidic technology to prepare alginate microspheres with different elasticities and architectures which they used as the carrier of MSCs. The elasticity of alginate hydrogels increased with increased calcium ion concentration, while the porosity decreased. MSCs can be encapsulated into these microspheres uniformly, and be regulated by the interplaying effect of elasticity and microstructure. Enzyme-catalysed crosslinking of silk fibroin using horseradish peroxidase and hydrogen peroxide has been reported to form hydrogels with adjustable stiffness in the range of 0.2–10 kPa. Wang et al.35 recently introduced inert silk fibroin nanofibres into these gels, and significantly improved the elasticity up to 9–60 kPa to match the mechanical properties of many tissues, especially bone and cartilage. To mimic the gradual change in elasticity of the annulus fibrosus (AF) along the radial direction, we synthesized biodegradable poly(ether carbonate urethane)urea with diverse Young’s moduli (2.5–13.4 MPa), by adjusting the molecular weight of polydiols and the feed ratios of hard molecular segment to soft molecular segment.36 ...
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Hydrogel elasticity and microarchitecture regulate dental-derived mesenchymal stem cell-host immune system cross-talk
2
2017
... Many natural or synthetic biomaterials with adjustable elasticity have been developed for cell modulation.20-33 In order to expand the library of controllable elastic biomaterials, study the interplaying effect of matrix elasticity with other physical cues, and develop elastic materials with tissue heterogeneity, some new strategies have been developed recently. For example, to understand the cross-talk effect of porosity and elasticity on encapsulated mesenchymal stem cells (MSCs), Ansari et al.34 used microfluidic technology to prepare alginate microspheres with different elasticities and architectures which they used as the carrier of MSCs. The elasticity of alginate hydrogels increased with increased calcium ion concentration, while the porosity decreased. MSCs can be encapsulated into these microspheres uniformly, and be regulated by the interplaying effect of elasticity and microstructure. Enzyme-catalysed crosslinking of silk fibroin using horseradish peroxidase and hydrogen peroxide has been reported to form hydrogels with adjustable stiffness in the range of 0.2–10 kPa. Wang et al.35 recently introduced inert silk fibroin nanofibres into these gels, and significantly improved the elasticity up to 9–60 kPa to match the mechanical properties of many tissues, especially bone and cartilage. To mimic the gradual change in elasticity of the annulus fibrosus (AF) along the radial direction, we synthesized biodegradable poly(ether carbonate urethane)urea with diverse Young’s moduli (2.5–13.4 MPa), by adjusting the molecular weight of polydiols and the feed ratios of hard molecular segment to soft molecular segment.36 ...
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Bioactive silk hydrogels with tunable mechanical properties
2
2018
... Many natural or synthetic biomaterials with adjustable elasticity have been developed for cell modulation.20-33 In order to expand the library of controllable elastic biomaterials, study the interplaying effect of matrix elasticity with other physical cues, and develop elastic materials with tissue heterogeneity, some new strategies have been developed recently. For example, to understand the cross-talk effect of porosity and elasticity on encapsulated mesenchymal stem cells (MSCs), Ansari et al.34 used microfluidic technology to prepare alginate microspheres with different elasticities and architectures which they used as the carrier of MSCs. The elasticity of alginate hydrogels increased with increased calcium ion concentration, while the porosity decreased. MSCs can be encapsulated into these microspheres uniformly, and be regulated by the interplaying effect of elasticity and microstructure. Enzyme-catalysed crosslinking of silk fibroin using horseradish peroxidase and hydrogen peroxide has been reported to form hydrogels with adjustable stiffness in the range of 0.2–10 kPa. Wang et al.35 recently introduced inert silk fibroin nanofibres into these gels, and significantly improved the elasticity up to 9–60 kPa to match the mechanical properties of many tissues, especially bone and cartilage. To mimic the gradual change in elasticity of the annulus fibrosus (AF) along the radial direction, we synthesized biodegradable poly(ether carbonate urethane)urea with diverse Young’s moduli (2.5–13.4 MPa), by adjusting the molecular weight of polydiols and the feed ratios of hard molecular segment to soft molecular segment.36 ...
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Modulation of the gene expression of annulus fibrosus-derived stem cells using poly(ether carbonate urethane)urea scaffolds of tunable elasticity
3
2016
... Many natural or synthetic biomaterials with adjustable elasticity have been developed for cell modulation.20-33 In order to expand the library of controllable elastic biomaterials, study the interplaying effect of matrix elasticity with other physical cues, and develop elastic materials with tissue heterogeneity, some new strategies have been developed recently. For example, to understand the cross-talk effect of porosity and elasticity on encapsulated mesenchymal stem cells (MSCs), Ansari et al.34 used microfluidic technology to prepare alginate microspheres with different elasticities and architectures which they used as the carrier of MSCs. The elasticity of alginate hydrogels increased with increased calcium ion concentration, while the porosity decreased. MSCs can be encapsulated into these microspheres uniformly, and be regulated by the interplaying effect of elasticity and microstructure. Enzyme-catalysed crosslinking of silk fibroin using horseradish peroxidase and hydrogen peroxide has been reported to form hydrogels with adjustable stiffness in the range of 0.2–10 kPa. Wang et al.35 recently introduced inert silk fibroin nanofibres into these gels, and significantly improved the elasticity up to 9–60 kPa to match the mechanical properties of many tissues, especially bone and cartilage. To mimic the gradual change in elasticity of the annulus fibrosus (AF) along the radial direction, we synthesized biodegradable poly(ether carbonate urethane)urea with diverse Young’s moduli (2.5–13.4 MPa), by adjusting the molecular weight of polydiols and the feed ratios of hard molecular segment to soft molecular segment.36 ...
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
PEG-anthracene hydrogels as an on-demand stiffening matrix to study mechanobiology
1
2019
... In natural tissues, ECM remodelling often changes the physical stiffness of the local environment. In the past, most research in this field was carried out using static substrates as artificial ECM. In recent years, some hydrogels with dynamic stiffness in situ have emerged to simulate the dynamic stiffness changes which occur in vivo. For example, Günay et al.37 used light-mediated anthracene group [4+4] photodimerization to prepare polyethylene glycol (PEG)-based hydrogels, which were cytocompatible and provided a tailorable initial modulus that could be further strengthened. To correspond to the stiffness of a healthy or a fibrotic heart, the hydrogels were stiffened from 10 kPa to 50 kPa to monitor the stiffness-dependent localization of nuclear factor of activated T cells. This type of material provides an intriguing way to reveal the dynamics of mechano-reactive signalling pathways, which differ from those under static cell culture. Moreover, Young and Engler38 prepared hydrogels with time-dependent stiffness by polymerization of thiolated hyaluronic acid and polyethylene glycol diacrylate (PEGDA) to simulate the dynamic mechanics of heart development. The stiffness of the hydrogel increased from 1.9 ± 0.1 kPa to 8.2 ± 1.1 kPa within 69.6 hours after polymerization, and ester hydrolysis had no significant effect on hydrogel stiffening over 2 weeks. Compared with the static polyacrylamide hydrogel group, pre-cardiac cells cultured on dynamic hyaluronic acid hydrogels formed more mature myocardial fibres and the expression level of mature cardiac-specific markers was significantly increased. More interestingly, Fu et al.39 used a redox-responsive protein folding switch to fabricate a dynamic protein hydrogel with a Young’s modulus which could be reversibly regulated using redox conditions. Whether starting from the reduced state or from the oxidized state, the Young’s modulus of the hydrogels was reversible cyclically in response to changes in the redox condition. Importantly, such reversibility remained even after storage in phosphate-buffered saline or cell culture medium for 2 weeks. They further found that human lung fibroblasts dynamically changed their morphology with the cyclic variation of the hydrogel’s Young’s modulus between ~6 kPa and ~20 kPa, demonstrating the potential of such material in mechanobiology studies. ...
Hydrogels with time-dependent material properties enhance cardiomyocyte differentiation in vitro
1
2011
... In natural tissues, ECM remodelling often changes the physical stiffness of the local environment. In the past, most research in this field was carried out using static substrates as artificial ECM. In recent years, some hydrogels with dynamic stiffness in situ have emerged to simulate the dynamic stiffness changes which occur in vivo. For example, Günay et al.37 used light-mediated anthracene group [4+4] photodimerization to prepare polyethylene glycol (PEG)-based hydrogels, which were cytocompatible and provided a tailorable initial modulus that could be further strengthened. To correspond to the stiffness of a healthy or a fibrotic heart, the hydrogels were stiffened from 10 kPa to 50 kPa to monitor the stiffness-dependent localization of nuclear factor of activated T cells. This type of material provides an intriguing way to reveal the dynamics of mechano-reactive signalling pathways, which differ from those under static cell culture. Moreover, Young and Engler38 prepared hydrogels with time-dependent stiffness by polymerization of thiolated hyaluronic acid and polyethylene glycol diacrylate (PEGDA) to simulate the dynamic mechanics of heart development. The stiffness of the hydrogel increased from 1.9 ± 0.1 kPa to 8.2 ± 1.1 kPa within 69.6 hours after polymerization, and ester hydrolysis had no significant effect on hydrogel stiffening over 2 weeks. Compared with the static polyacrylamide hydrogel group, pre-cardiac cells cultured on dynamic hyaluronic acid hydrogels formed more mature myocardial fibres and the expression level of mature cardiac-specific markers was significantly increased. More interestingly, Fu et al.39 used a redox-responsive protein folding switch to fabricate a dynamic protein hydrogel with a Young’s modulus which could be reversibly regulated using redox conditions. Whether starting from the reduced state or from the oxidized state, the Young’s modulus of the hydrogels was reversible cyclically in response to changes in the redox condition. Importantly, such reversibility remained even after storage in phosphate-buffered saline or cell culture medium for 2 weeks. They further found that human lung fibroblasts dynamically changed their morphology with the cyclic variation of the hydrogel’s Young’s modulus between ~6 kPa and ~20 kPa, demonstrating the potential of such material in mechanobiology studies. ...
Dynamic protein hydrogels with reversibly tunable stiffness regulate human lung fibroblast spreading reversibly
3
2019
... In natural tissues, ECM remodelling often changes the physical stiffness of the local environment. In the past, most research in this field was carried out using static substrates as artificial ECM. In recent years, some hydrogels with dynamic stiffness in situ have emerged to simulate the dynamic stiffness changes which occur in vivo. For example, Günay et al.37 used light-mediated anthracene group [4+4] photodimerization to prepare polyethylene glycol (PEG)-based hydrogels, which were cytocompatible and provided a tailorable initial modulus that could be further strengthened. To correspond to the stiffness of a healthy or a fibrotic heart, the hydrogels were stiffened from 10 kPa to 50 kPa to monitor the stiffness-dependent localization of nuclear factor of activated T cells. This type of material provides an intriguing way to reveal the dynamics of mechano-reactive signalling pathways, which differ from those under static cell culture. Moreover, Young and Engler38 prepared hydrogels with time-dependent stiffness by polymerization of thiolated hyaluronic acid and polyethylene glycol diacrylate (PEGDA) to simulate the dynamic mechanics of heart development. The stiffness of the hydrogel increased from 1.9 ± 0.1 kPa to 8.2 ± 1.1 kPa within 69.6 hours after polymerization, and ester hydrolysis had no significant effect on hydrogel stiffening over 2 weeks. Compared with the static polyacrylamide hydrogel group, pre-cardiac cells cultured on dynamic hyaluronic acid hydrogels formed more mature myocardial fibres and the expression level of mature cardiac-specific markers was significantly increased. More interestingly, Fu et al.39 used a redox-responsive protein folding switch to fabricate a dynamic protein hydrogel with a Young’s modulus which could be reversibly regulated using redox conditions. Whether starting from the reduced state or from the oxidized state, the Young’s modulus of the hydrogels was reversible cyclically in response to changes in the redox condition. Importantly, such reversibility remained even after storage in phosphate-buffered saline or cell culture medium for 2 weeks. They further found that human lung fibroblasts dynamically changed their morphology with the cyclic variation of the hydrogel’s Young’s modulus between ~6 kPa and ~20 kPa, demonstrating the potential of such material in mechanobiology studies. ...
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Extracellular matrix elasticity regulates osteocyte gap junction elongation: involvement of paxillin in intracellular signal transduction
1
2018
... Cell adhesion to the ECM is a complex process, involving cell attachment, cell spreading, actin cytoskeleton organization and then focal adhesion (FA) formation. Cell attachment can be completed in a short time to connect cells to the matrix. The cells then spread on the matrix and develop different morphologies. Following that, actin polymerises into microfilaments to form the actin cytoskeleton. Finally, the formation of FAs constitutes the connection between the matrix and the cellular cytoskeleton. The FAs, mainly composed of transmembrane adhesion receptors including integrins, vinculins and paxillins, can transfer biological and mechanical stimuli into cells and initiate molecular pathways which subsequently regulate gene expression.20 A number of studies have demonstrated that the assembly and turnover of FAs are affected by the elasticity of the substrate. In general, cells have more stable and matured FAs on stiffer substrates, and have reduced and diffused adhesion complexes on softer substrates. For example, osteocytes form more stable vinculin-containing adhesion complexes on a stiff substrate than those on a soft substrate.40 Myocardial cells also exhibit a stronger traction force on stiffer substrates.41 However, the relationship between cell adhesion strength and matrix elasticity is non-linear. It has been reported that when fibroblasts and endothelial cells (ECs) are cultured sparsely without cell-cell contacts, the spread area of cells abruptly changes at a stiffness range of around 3 kPa.42 ...
Substrate stiffness-regulated matrix metalloproteinase output in myocardial cells and cardiac fibroblasts: implications for myocardial fibrosis
1
2014
... Cell adhesion to the ECM is a complex process, involving cell attachment, cell spreading, actin cytoskeleton organization and then focal adhesion (FA) formation. Cell attachment can be completed in a short time to connect cells to the matrix. The cells then spread on the matrix and develop different morphologies. Following that, actin polymerises into microfilaments to form the actin cytoskeleton. Finally, the formation of FAs constitutes the connection between the matrix and the cellular cytoskeleton. The FAs, mainly composed of transmembrane adhesion receptors including integrins, vinculins and paxillins, can transfer biological and mechanical stimuli into cells and initiate molecular pathways which subsequently regulate gene expression.20 A number of studies have demonstrated that the assembly and turnover of FAs are affected by the elasticity of the substrate. In general, cells have more stable and matured FAs on stiffer substrates, and have reduced and diffused adhesion complexes on softer substrates. For example, osteocytes form more stable vinculin-containing adhesion complexes on a stiff substrate than those on a soft substrate.40 Myocardial cells also exhibit a stronger traction force on stiffer substrates.41 However, the relationship between cell adhesion strength and matrix elasticity is non-linear. It has been reported that when fibroblasts and endothelial cells (ECs) are cultured sparsely without cell-cell contacts, the spread area of cells abruptly changes at a stiffness range of around 3 kPa.42 ...
Effects of substrate stiffness on cell morphology, cytoskeletal structure, and adhesion
1
2005
... Cell adhesion to the ECM is a complex process, involving cell attachment, cell spreading, actin cytoskeleton organization and then focal adhesion (FA) formation. Cell attachment can be completed in a short time to connect cells to the matrix. The cells then spread on the matrix and develop different morphologies. Following that, actin polymerises into microfilaments to form the actin cytoskeleton. Finally, the formation of FAs constitutes the connection between the matrix and the cellular cytoskeleton. The FAs, mainly composed of transmembrane adhesion receptors including integrins, vinculins and paxillins, can transfer biological and mechanical stimuli into cells and initiate molecular pathways which subsequently regulate gene expression.20 A number of studies have demonstrated that the assembly and turnover of FAs are affected by the elasticity of the substrate. In general, cells have more stable and matured FAs on stiffer substrates, and have reduced and diffused adhesion complexes on softer substrates. For example, osteocytes form more stable vinculin-containing adhesion complexes on a stiff substrate than those on a soft substrate.40 Myocardial cells also exhibit a stronger traction force on stiffer substrates.41 However, the relationship between cell adhesion strength and matrix elasticity is non-linear. It has been reported that when fibroblasts and endothelial cells (ECs) are cultured sparsely without cell-cell contacts, the spread area of cells abruptly changes at a stiffness range of around 3 kPa.42 ...
Biomaterial arrays with defined adhesion ligand densities and matrix stiffness identify distinct phenotypes for tumorigenic and nontumorigenic human mesenchymal cell types
1
2014
... Cell migration is an essential cell behaviour in diverse biological processes including development, angiogenesis, wound healing and inflammation. It is also highly sensitive to environmental stiffness. For example, the migration speed of HT-1080 fibrosarcoma cells doubled on a stiff gel (13 kPa) compared to those on a soft gel (0.34 kPa).43 When renal progenitor cells were seeded onto substrates with different elasticities, cell migration was limited on the substrates of 0.5 kPa and 2 kPa, but increased on the substrate of 4 kPa and remained stable on the stiff plates.44 Migration speed thus increases or decreases with the change of matrix elasticity, in other words, many cell types have an optimal stiffness, at which cell migration is maximal. For instance, smooth muscle cells show obvious biphasic dependence on matrix elasticity, and have a maximal speed at a particular stiffness.45 To predict the optimal environmental stiffness for different cells, Bangasserere et al.46 developed a cell migration simulator and successfully determined the optimal stiffness for embryonic chick forebrain neurons (~1 kPa) and U251 glioma cells (~100 kPa). The simulator would help researchers find the optimum stiffness to encourage cells to move to damaged or unhealthy tissues to restore the function of the tissue. Matrix elasticity affects not only the ability of cells to migrate, but also their direction of migration. Directional migration has been observed from softer to more stiff regions for many cell types.47-49 For example, vascular smooth muscle cells showed obvious directional migration on a mechanical gradient matrix coated with fibronectin, but not on a mechanical gradient matrix coated with laminin, indicating that the adhesion ligand might be an important element which affects cell migration in response to mechanical gradients.47 ...
Substrate stiffness modulates renal progenitor cell properties via a ROCK-mediated mechanotransduction Mechanism
1
2019
... Cell migration is an essential cell behaviour in diverse biological processes including development, angiogenesis, wound healing and inflammation. It is also highly sensitive to environmental stiffness. For example, the migration speed of HT-1080 fibrosarcoma cells doubled on a stiff gel (13 kPa) compared to those on a soft gel (0.34 kPa).43 When renal progenitor cells were seeded onto substrates with different elasticities, cell migration was limited on the substrates of 0.5 kPa and 2 kPa, but increased on the substrate of 4 kPa and remained stable on the stiff plates.44 Migration speed thus increases or decreases with the change of matrix elasticity, in other words, many cell types have an optimal stiffness, at which cell migration is maximal. For instance, smooth muscle cells show obvious biphasic dependence on matrix elasticity, and have a maximal speed at a particular stiffness.45 To predict the optimal environmental stiffness for different cells, Bangasserere et al.46 developed a cell migration simulator and successfully determined the optimal stiffness for embryonic chick forebrain neurons (~1 kPa) and U251 glioma cells (~100 kPa). The simulator would help researchers find the optimum stiffness to encourage cells to move to damaged or unhealthy tissues to restore the function of the tissue. Matrix elasticity affects not only the ability of cells to migrate, but also their direction of migration. Directional migration has been observed from softer to more stiff regions for many cell types.47-49 For example, vascular smooth muscle cells showed obvious directional migration on a mechanical gradient matrix coated with fibronectin, but not on a mechanical gradient matrix coated with laminin, indicating that the adhesion ligand might be an important element which affects cell migration in response to mechanical gradients.47 ...
Extracellular matrix rigidity governs smooth muscle cell motility in a biphasic fashion
1
2005
... Cell migration is an essential cell behaviour in diverse biological processes including development, angiogenesis, wound healing and inflammation. It is also highly sensitive to environmental stiffness. For example, the migration speed of HT-1080 fibrosarcoma cells doubled on a stiff gel (13 kPa) compared to those on a soft gel (0.34 kPa).43 When renal progenitor cells were seeded onto substrates with different elasticities, cell migration was limited on the substrates of 0.5 kPa and 2 kPa, but increased on the substrate of 4 kPa and remained stable on the stiff plates.44 Migration speed thus increases or decreases with the change of matrix elasticity, in other words, many cell types have an optimal stiffness, at which cell migration is maximal. For instance, smooth muscle cells show obvious biphasic dependence on matrix elasticity, and have a maximal speed at a particular stiffness.45 To predict the optimal environmental stiffness for different cells, Bangasserere et al.46 developed a cell migration simulator and successfully determined the optimal stiffness for embryonic chick forebrain neurons (~1 kPa) and U251 glioma cells (~100 kPa). The simulator would help researchers find the optimum stiffness to encourage cells to move to damaged or unhealthy tissues to restore the function of the tissue. Matrix elasticity affects not only the ability of cells to migrate, but also their direction of migration. Directional migration has been observed from softer to more stiff regions for many cell types.47-49 For example, vascular smooth muscle cells showed obvious directional migration on a mechanical gradient matrix coated with fibronectin, but not on a mechanical gradient matrix coated with laminin, indicating that the adhesion ligand might be an important element which affects cell migration in response to mechanical gradients.47 ...
Shifting the optimal stiffness for cell migration
1
2017
... Cell migration is an essential cell behaviour in diverse biological processes including development, angiogenesis, wound healing and inflammation. It is also highly sensitive to environmental stiffness. For example, the migration speed of HT-1080 fibrosarcoma cells doubled on a stiff gel (13 kPa) compared to those on a soft gel (0.34 kPa).43 When renal progenitor cells were seeded onto substrates with different elasticities, cell migration was limited on the substrates of 0.5 kPa and 2 kPa, but increased on the substrate of 4 kPa and remained stable on the stiff plates.44 Migration speed thus increases or decreases with the change of matrix elasticity, in other words, many cell types have an optimal stiffness, at which cell migration is maximal. For instance, smooth muscle cells show obvious biphasic dependence on matrix elasticity, and have a maximal speed at a particular stiffness.45 To predict the optimal environmental stiffness for different cells, Bangasserere et al.46 developed a cell migration simulator and successfully determined the optimal stiffness for embryonic chick forebrain neurons (~1 kPa) and U251 glioma cells (~100 kPa). The simulator would help researchers find the optimum stiffness to encourage cells to move to damaged or unhealthy tissues to restore the function of the tissue. Matrix elasticity affects not only the ability of cells to migrate, but also their direction of migration. Directional migration has been observed from softer to more stiff regions for many cell types.47-49 For example, vascular smooth muscle cells showed obvious directional migration on a mechanical gradient matrix coated with fibronectin, but not on a mechanical gradient matrix coated with laminin, indicating that the adhesion ligand might be an important element which affects cell migration in response to mechanical gradients.47 ...
Cell movement is guided by the rigidity of the substrate
2
2000
... Cell migration is an essential cell behaviour in diverse biological processes including development, angiogenesis, wound healing and inflammation. It is also highly sensitive to environmental stiffness. For example, the migration speed of HT-1080 fibrosarcoma cells doubled on a stiff gel (13 kPa) compared to those on a soft gel (0.34 kPa).43 When renal progenitor cells were seeded onto substrates with different elasticities, cell migration was limited on the substrates of 0.5 kPa and 2 kPa, but increased on the substrate of 4 kPa and remained stable on the stiff plates.44 Migration speed thus increases or decreases with the change of matrix elasticity, in other words, many cell types have an optimal stiffness, at which cell migration is maximal. For instance, smooth muscle cells show obvious biphasic dependence on matrix elasticity, and have a maximal speed at a particular stiffness.45 To predict the optimal environmental stiffness for different cells, Bangasserere et al.46 developed a cell migration simulator and successfully determined the optimal stiffness for embryonic chick forebrain neurons (~1 kPa) and U251 glioma cells (~100 kPa). The simulator would help researchers find the optimum stiffness to encourage cells to move to damaged or unhealthy tissues to restore the function of the tissue. Matrix elasticity affects not only the ability of cells to migrate, but also their direction of migration. Directional migration has been observed from softer to more stiff regions for many cell types.47-49 For example, vascular smooth muscle cells showed obvious directional migration on a mechanical gradient matrix coated with fibronectin, but not on a mechanical gradient matrix coated with laminin, indicating that the adhesion ligand might be an important element which affects cell migration in response to mechanical gradients.47 ...
... 47 ...
Sensing of substratum rigidity and directional migration by fast-crawling cells
0
2018
Vascular smooth muscle cell durotaxis depends on extracellular matrix composition
1
2016
... Cell migration is an essential cell behaviour in diverse biological processes including development, angiogenesis, wound healing and inflammation. It is also highly sensitive to environmental stiffness. For example, the migration speed of HT-1080 fibrosarcoma cells doubled on a stiff gel (13 kPa) compared to those on a soft gel (0.34 kPa).43 When renal progenitor cells were seeded onto substrates with different elasticities, cell migration was limited on the substrates of 0.5 kPa and 2 kPa, but increased on the substrate of 4 kPa and remained stable on the stiff plates.44 Migration speed thus increases or decreases with the change of matrix elasticity, in other words, many cell types have an optimal stiffness, at which cell migration is maximal. For instance, smooth muscle cells show obvious biphasic dependence on matrix elasticity, and have a maximal speed at a particular stiffness.45 To predict the optimal environmental stiffness for different cells, Bangasserere et al.46 developed a cell migration simulator and successfully determined the optimal stiffness for embryonic chick forebrain neurons (~1 kPa) and U251 glioma cells (~100 kPa). The simulator would help researchers find the optimum stiffness to encourage cells to move to damaged or unhealthy tissues to restore the function of the tissue. Matrix elasticity affects not only the ability of cells to migrate, but also their direction of migration. Directional migration has been observed from softer to more stiff regions for many cell types.47-49 For example, vascular smooth muscle cells showed obvious directional migration on a mechanical gradient matrix coated with fibronectin, but not on a mechanical gradient matrix coated with laminin, indicating that the adhesion ligand might be an important element which affects cell migration in response to mechanical gradients.47 ...
Effect of substrate stiffness on proliferation and differentiation of periodontal ligament stem cells
1
2018
... Cell proliferation and apoptosis is the basis of growth, development and regeneration of organisms. There are many important factors that affect cell proliferation and apoptosis, and matrix elasticity is among them. For example, proliferation of periodontal ligament stem cells on stiff substrates was faster than on soft substrates.50 The effect of matrix stiffness on cell proliferation is especially prominent in tumour cells. Increased proliferation of breast cell lines has been seen on stiff substrates.51 Many cells divide much more rapidly on a relatively stiffer substrate, while some cells behave differently. For instance, neural stem/progenitor cells proliferated well on soft substrates (< 10 kPa) and exhibited maximal proliferation on 3.5 kPa substrates.52 Robinson et al.53 developed a series of PEG-based hydrogels to investigate the effect of substrate stiffness on various vascular cells, including human umbilical vein ECs, human aortic adventitial fibroblasts, and human aortic vascular smooth muscle cells. Proliferation of human aortic adventitial fibroblasts increased with the increased hydrogel elasticity, yet proliferation of human umbilical vein ECs demonstrated an opposite trend with most rapid proliferation on the softest hydrogels. Interestingly, proliferation of human aortic vascular smooth muscle cells was hardly affected by matrix stiffness. Therefore, the proliferative response of cells to matrix elasticity showed cell-type specificity and nonlinearity. Accordingly, a range of substrate stiffness should be tested to determine the optimum moduli for cell growth. ECM with appropriate stiffness may promote cell proliferation and slow apoptosis. ...
Behavioral remodeling of normal and cancerous epithelial cell lines with differing invasion potential induced by substrate elastic modulus
1
2018
... Cell proliferation and apoptosis is the basis of growth, development and regeneration of organisms. There are many important factors that affect cell proliferation and apoptosis, and matrix elasticity is among them. For example, proliferation of periodontal ligament stem cells on stiff substrates was faster than on soft substrates.50 The effect of matrix stiffness on cell proliferation is especially prominent in tumour cells. Increased proliferation of breast cell lines has been seen on stiff substrates.51 Many cells divide much more rapidly on a relatively stiffer substrate, while some cells behave differently. For instance, neural stem/progenitor cells proliferated well on soft substrates (< 10 kPa) and exhibited maximal proliferation on 3.5 kPa substrates.52 Robinson et al.53 developed a series of PEG-based hydrogels to investigate the effect of substrate stiffness on various vascular cells, including human umbilical vein ECs, human aortic adventitial fibroblasts, and human aortic vascular smooth muscle cells. Proliferation of human aortic adventitial fibroblasts increased with the increased hydrogel elasticity, yet proliferation of human umbilical vein ECs demonstrated an opposite trend with most rapid proliferation on the softest hydrogels. Interestingly, proliferation of human aortic vascular smooth muscle cells was hardly affected by matrix stiffness. Therefore, the proliferative response of cells to matrix elasticity showed cell-type specificity and nonlinearity. Accordingly, a range of substrate stiffness should be tested to determine the optimum moduli for cell growth. ECM with appropriate stiffness may promote cell proliferation and slow apoptosis. ...
The effect of substrate stiffness on adult neural stem cell behavior
1
2009
... Cell proliferation and apoptosis is the basis of growth, development and regeneration of organisms. There are many important factors that affect cell proliferation and apoptosis, and matrix elasticity is among them. For example, proliferation of periodontal ligament stem cells on stiff substrates was faster than on soft substrates.50 The effect of matrix stiffness on cell proliferation is especially prominent in tumour cells. Increased proliferation of breast cell lines has been seen on stiff substrates.51 Many cells divide much more rapidly on a relatively stiffer substrate, while some cells behave differently. For instance, neural stem/progenitor cells proliferated well on soft substrates (< 10 kPa) and exhibited maximal proliferation on 3.5 kPa substrates.52 Robinson et al.53 developed a series of PEG-based hydrogels to investigate the effect of substrate stiffness on various vascular cells, including human umbilical vein ECs, human aortic adventitial fibroblasts, and human aortic vascular smooth muscle cells. Proliferation of human aortic adventitial fibroblasts increased with the increased hydrogel elasticity, yet proliferation of human umbilical vein ECs demonstrated an opposite trend with most rapid proliferation on the softest hydrogels. Interestingly, proliferation of human aortic vascular smooth muscle cells was hardly affected by matrix stiffness. Therefore, the proliferative response of cells to matrix elasticity showed cell-type specificity and nonlinearity. Accordingly, a range of substrate stiffness should be tested to determine the optimum moduli for cell growth. ECM with appropriate stiffness may promote cell proliferation and slow apoptosis. ...
Differential effects of substrate modulus on human vascular endothelial, smooth muscle, and fibroblastic cells
1
2012
... Cell proliferation and apoptosis is the basis of growth, development and regeneration of organisms. There are many important factors that affect cell proliferation and apoptosis, and matrix elasticity is among them. For example, proliferation of periodontal ligament stem cells on stiff substrates was faster than on soft substrates.50 The effect of matrix stiffness on cell proliferation is especially prominent in tumour cells. Increased proliferation of breast cell lines has been seen on stiff substrates.51 Many cells divide much more rapidly on a relatively stiffer substrate, while some cells behave differently. For instance, neural stem/progenitor cells proliferated well on soft substrates (< 10 kPa) and exhibited maximal proliferation on 3.5 kPa substrates.52 Robinson et al.53 developed a series of PEG-based hydrogels to investigate the effect of substrate stiffness on various vascular cells, including human umbilical vein ECs, human aortic adventitial fibroblasts, and human aortic vascular smooth muscle cells. Proliferation of human aortic adventitial fibroblasts increased with the increased hydrogel elasticity, yet proliferation of human umbilical vein ECs demonstrated an opposite trend with most rapid proliferation on the softest hydrogels. Interestingly, proliferation of human aortic vascular smooth muscle cells was hardly affected by matrix stiffness. Therefore, the proliferative response of cells to matrix elasticity showed cell-type specificity and nonlinearity. Accordingly, a range of substrate stiffness should be tested to determine the optimum moduli for cell growth. ECM with appropriate stiffness may promote cell proliferation and slow apoptosis. ...
Effect of substrate stiffness on early human embryonic stem cell differentiation
1
2013
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
Rigid microenvironments promote cardiac differentiation of mouse and human embryonic stem cells
1
2013
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
Substrate stiffness and composition specifically direct differentiation of induced pluripotent stem cells
1
2015
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
Respective effects of gelatin-coated polydimethylsiloxane (PDMS) substrates on self-renewal and cardiac differentiation of induced pluripotent stem cells (iPSCs)
2
2018
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
... 57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
Stiffness sensing by cells
1
2020
... Stem cells can self-renew and differentiate, playing an important role in tissue regeneration. Matrix elasticity has been considered to be one of the key factors that regulate the fate of stem cells, including embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and various adult stem cells. For example, when human ESCs were cultured on fibronectin-coated poly(dimethyl siloxane) (PDMS) substrates with various elasticities,54 mesodermal differentiation was upregulated as stiffness increased. In another study, Arshi et al.55 studied the effect of matrix rigidity on cardiac cell differentiation by using mouse and human ESCs. They found that a rigid microenvironment promoted cardiac differentiation of both mouse and human ESCs. Similar to ESCs, iPSCs can also differentiate into cardiomyocytes on soft polyacrylamide gels.56 Recently, Fu et al.57 utilised gelatine-coated PDMS substrates to study the influence of stiffness on the self-renewal and cardiac differentiation of iPSCs. They found that iPSCs exhibited higher levels of a pluripotency marker, but lower cardiac gene expression, on stiffer PDMS, while iPSCs on soft PDMS responded in the opposite way.57 MSCs are a promising adult stem cell source for tissue regeneration, which have been more commonly used in studying the role of matrix elasticity on stem-cell fate. For example, Discher et al.20 demonstrated that human MSCs (hMSCs) differentiated into neuron-like cells, myoblasts, and osteoblasts, on gels whose elasticity mimicked the corresponding tissues. Subsequently, extensive studies have confirmed the relationship between matrix elasticity and MSC differentiation, including osteogenesis, myogenesis, adipogenesis, angiogenesis, and neurogenesis. For instance, MSCs undergo stronger chondrogenic differentiation on stiff matrix compared with soft matrix.39 Thus tissue-derived stem cells can be easily induced into the corresponding tissues. In order to check whether AF-derived stem cells (AFSCs) can differentiate into corresponding cell types in native AF tissue to produce matching ECM, we recently developed a series of polycarbonate-based polyurethanes to mimic the physical features of natural AF tissue. We found that the elasticity of the scaffold has the ability to induce AFSCs to differentiate into corresponding AF-like cells.36 In general, stem cells differentiate best on substrates which have similar elasticity to the target tissue.58 ...
Matrix stiffness modulates formation and activity of neuronal networks of controlled architectures
1
2016
... Matrix elasticity has a great impact on cell fate and activity. Therefore, this feature must be considered carefully when developing biomaterials for tissue engineering. Choosing suitable matrix stiffness to promote tissue regeneration has been widely used in the regeneration of tissues including nerve, muscle, bone, cartilage, and blood vessels. For example, the formation and activity of neuronal networks can be modulated by matrix stiffness.59 On soft substrates, the migration of cortical neurons is enhanced, which leads to a faster formation of neuronal networks, while on stiff substrates, the pre-synaptic density, number of action potentials and miniature synaptic currents are enhanced, which indicates that matrix stiffness should be a key design parameter in scaffold fabrication for neural tissue engineering. However, for bone tissue engineering, a stiffer substrate is more appropriate. For example, three-dimensional (3D) porous collagen/hydroxyapatite scaffolds with the same microstructures but different stiffnesses were fabricated for bone tissue engineering.60 Bone matrix proteins, including osteocalcin and osteopontin, exhibited significantly higher expression on the highest stiffness group after 3 weeks of culture. The expression of osteocalcin and osteopontin was also higher on the highest stiffness group after 1 and 3 months of implantation in vivo. Matrix elasticity also plays a critical role in regulating chondrocyte behaviour and phenotype in vitro for cartilage tissue engineering. Many key features of chondrocytes including cell spreading area, percentage of irregular cell shape distribution, and mechanical characteristics are matrix stiffness-dependent,61 implying that choosing the optimal matrix elasticity is very important for cartilage tissue engineering.62-64 For instance, architectured 3D hydrogel scaffolds (ArcGels) were used to explore the stiffness effect on human articular chondrocytes in vitro and in vivo. The results showed that chondrogenic differentiation was more efficiently promoted on soft ArcGel in vitro, while the chondrogenic phenotype of human articular chondrocytes and retention of cartilaginous matrix were better maintained on stiff ArcGel upon ectopic implantation. In general, when the stiffness of a tissue engineering scaffold is similar to that of the host tissue, the stem cells tend to proliferate, secrete a large amount of ECM and differentiate into the cells of the specific tissue. ...
3D scaffolds with different stiffness but the same microstructure for bone tissue engineering
1
2015
... Matrix elasticity has a great impact on cell fate and activity. Therefore, this feature must be considered carefully when developing biomaterials for tissue engineering. Choosing suitable matrix stiffness to promote tissue regeneration has been widely used in the regeneration of tissues including nerve, muscle, bone, cartilage, and blood vessels. For example, the formation and activity of neuronal networks can be modulated by matrix stiffness.59 On soft substrates, the migration of cortical neurons is enhanced, which leads to a faster formation of neuronal networks, while on stiff substrates, the pre-synaptic density, number of action potentials and miniature synaptic currents are enhanced, which indicates that matrix stiffness should be a key design parameter in scaffold fabrication for neural tissue engineering. However, for bone tissue engineering, a stiffer substrate is more appropriate. For example, three-dimensional (3D) porous collagen/hydroxyapatite scaffolds with the same microstructures but different stiffnesses were fabricated for bone tissue engineering.60 Bone matrix proteins, including osteocalcin and osteopontin, exhibited significantly higher expression on the highest stiffness group after 3 weeks of culture. The expression of osteocalcin and osteopontin was also higher on the highest stiffness group after 1 and 3 months of implantation in vivo. Matrix elasticity also plays a critical role in regulating chondrocyte behaviour and phenotype in vitro for cartilage tissue engineering. Many key features of chondrocytes including cell spreading area, percentage of irregular cell shape distribution, and mechanical characteristics are matrix stiffness-dependent,61 implying that choosing the optimal matrix elasticity is very important for cartilage tissue engineering.62-64 For instance, architectured 3D hydrogel scaffolds (ArcGels) were used to explore the stiffness effect on human articular chondrocytes in vitro and in vivo. The results showed that chondrogenic differentiation was more efficiently promoted on soft ArcGel in vitro, while the chondrogenic phenotype of human articular chondrocytes and retention of cartilaginous matrix were better maintained on stiff ArcGel upon ectopic implantation. In general, when the stiffness of a tissue engineering scaffold is similar to that of the host tissue, the stem cells tend to proliferate, secrete a large amount of ECM and differentiate into the cells of the specific tissue. ...
The effect of matrix stiffness on biomechanical properties of chondrocytes
1
2016
... Matrix elasticity has a great impact on cell fate and activity. Therefore, this feature must be considered carefully when developing biomaterials for tissue engineering. Choosing suitable matrix stiffness to promote tissue regeneration has been widely used in the regeneration of tissues including nerve, muscle, bone, cartilage, and blood vessels. For example, the formation and activity of neuronal networks can be modulated by matrix stiffness.59 On soft substrates, the migration of cortical neurons is enhanced, which leads to a faster formation of neuronal networks, while on stiff substrates, the pre-synaptic density, number of action potentials and miniature synaptic currents are enhanced, which indicates that matrix stiffness should be a key design parameter in scaffold fabrication for neural tissue engineering. However, for bone tissue engineering, a stiffer substrate is more appropriate. For example, three-dimensional (3D) porous collagen/hydroxyapatite scaffolds with the same microstructures but different stiffnesses were fabricated for bone tissue engineering.60 Bone matrix proteins, including osteocalcin and osteopontin, exhibited significantly higher expression on the highest stiffness group after 3 weeks of culture. The expression of osteocalcin and osteopontin was also higher on the highest stiffness group after 1 and 3 months of implantation in vivo. Matrix elasticity also plays a critical role in regulating chondrocyte behaviour and phenotype in vitro for cartilage tissue engineering. Many key features of chondrocytes including cell spreading area, percentage of irregular cell shape distribution, and mechanical characteristics are matrix stiffness-dependent,61 implying that choosing the optimal matrix elasticity is very important for cartilage tissue engineering.62-64 For instance, architectured 3D hydrogel scaffolds (ArcGels) were used to explore the stiffness effect on human articular chondrocytes in vitro and in vivo. The results showed that chondrogenic differentiation was more efficiently promoted on soft ArcGel in vitro, while the chondrogenic phenotype of human articular chondrocytes and retention of cartilaginous matrix were better maintained on stiff ArcGel upon ectopic implantation. In general, when the stiffness of a tissue engineering scaffold is similar to that of the host tissue, the stem cells tend to proliferate, secrete a large amount of ECM and differentiate into the cells of the specific tissue. ...
Chondrogenesis of human bone marrow mesenchymal stem cells in 3-dimensional, photocrosslinked hydrogel constructs: Effect of cell seeding density and material stiffness
1
2017
... Matrix elasticity has a great impact on cell fate and activity. Therefore, this feature must be considered carefully when developing biomaterials for tissue engineering. Choosing suitable matrix stiffness to promote tissue regeneration has been widely used in the regeneration of tissues including nerve, muscle, bone, cartilage, and blood vessels. For example, the formation and activity of neuronal networks can be modulated by matrix stiffness.59 On soft substrates, the migration of cortical neurons is enhanced, which leads to a faster formation of neuronal networks, while on stiff substrates, the pre-synaptic density, number of action potentials and miniature synaptic currents are enhanced, which indicates that matrix stiffness should be a key design parameter in scaffold fabrication for neural tissue engineering. However, for bone tissue engineering, a stiffer substrate is more appropriate. For example, three-dimensional (3D) porous collagen/hydroxyapatite scaffolds with the same microstructures but different stiffnesses were fabricated for bone tissue engineering.60 Bone matrix proteins, including osteocalcin and osteopontin, exhibited significantly higher expression on the highest stiffness group after 3 weeks of culture. The expression of osteocalcin and osteopontin was also higher on the highest stiffness group after 1 and 3 months of implantation in vivo. Matrix elasticity also plays a critical role in regulating chondrocyte behaviour and phenotype in vitro for cartilage tissue engineering. Many key features of chondrocytes including cell spreading area, percentage of irregular cell shape distribution, and mechanical characteristics are matrix stiffness-dependent,61 implying that choosing the optimal matrix elasticity is very important for cartilage tissue engineering.62-64 For instance, architectured 3D hydrogel scaffolds (ArcGels) were used to explore the stiffness effect on human articular chondrocytes in vitro and in vivo. The results showed that chondrogenic differentiation was more efficiently promoted on soft ArcGel in vitro, while the chondrogenic phenotype of human articular chondrocytes and retention of cartilaginous matrix were better maintained on stiff ArcGel upon ectopic implantation. In general, when the stiffness of a tissue engineering scaffold is similar to that of the host tissue, the stem cells tend to proliferate, secrete a large amount of ECM and differentiate into the cells of the specific tissue. ...
Interplay between stiffness and degradation of architectured gelatin hydrogels leads to differential modulation of chondrogenesis in vitro and in vivo
0
2018
Effect of matrix stiffness and adhesion ligand density on chondrogenic differentiation of mesenchymal stem cells
2
2020
... Matrix elasticity has a great impact on cell fate and activity. Therefore, this feature must be considered carefully when developing biomaterials for tissue engineering. Choosing suitable matrix stiffness to promote tissue regeneration has been widely used in the regeneration of tissues including nerve, muscle, bone, cartilage, and blood vessels. For example, the formation and activity of neuronal networks can be modulated by matrix stiffness.59 On soft substrates, the migration of cortical neurons is enhanced, which leads to a faster formation of neuronal networks, while on stiff substrates, the pre-synaptic density, number of action potentials and miniature synaptic currents are enhanced, which indicates that matrix stiffness should be a key design parameter in scaffold fabrication for neural tissue engineering. However, for bone tissue engineering, a stiffer substrate is more appropriate. For example, three-dimensional (3D) porous collagen/hydroxyapatite scaffolds with the same microstructures but different stiffnesses were fabricated for bone tissue engineering.60 Bone matrix proteins, including osteocalcin and osteopontin, exhibited significantly higher expression on the highest stiffness group after 3 weeks of culture. The expression of osteocalcin and osteopontin was also higher on the highest stiffness group after 1 and 3 months of implantation in vivo. Matrix elasticity also plays a critical role in regulating chondrocyte behaviour and phenotype in vitro for cartilage tissue engineering. Many key features of chondrocytes including cell spreading area, percentage of irregular cell shape distribution, and mechanical characteristics are matrix stiffness-dependent,61 implying that choosing the optimal matrix elasticity is very important for cartilage tissue engineering.62-64 For instance, architectured 3D hydrogel scaffolds (ArcGels) were used to explore the stiffness effect on human articular chondrocytes in vitro and in vivo. The results showed that chondrogenic differentiation was more efficiently promoted on soft ArcGel in vitro, while the chondrogenic phenotype of human articular chondrocytes and retention of cartilaginous matrix were better maintained on stiff ArcGel upon ectopic implantation. In general, when the stiffness of a tissue engineering scaffold is similar to that of the host tissue, the stem cells tend to proliferate, secrete a large amount of ECM and differentiate into the cells of the specific tissue. ...
... The proliferation of various resident cells in a viscoelastic microenvironment was also found to be enhanced and regulated. Several studies demonstrated that hydrogel viscoelasticity has an obvious effect in promoting spreading and proliferation of fibroblasts in 2D or 3D culture.73, 88, 89 Similarly, when hMSCs were cultured on polyacrylamide-based viscoelastic hydrogels, the proliferation rate of hMSCs was significantly promoted by faster stress relaxation times.64 On an oxime cross-linked alginate hydrogel,82 2PK3 cells showed the same trend, with cells growing better in hydrogels with fast stress relaxation. All results indicate that stress relaxation time-dependent regulation may have beneficial effects on proliferation. ...
The anisotropic mechanical behaviour of passive skeletal muscle tissue subjected to large tensile strain
1
2013
... Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation. ...
Assessment of liver viscoelasticity using multifrequency MR elastography
1
2008
... Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation. ...
Breast elasticity: principles, technique, results: an update and overview of commercially available software
0
2013
Dynamic viscoelastic models of human skin using optical elastography
2
2015
... The methods used to explore the molecular mechanism of how cells sense ECM viscoelasticity are similar to those used in studying how cells sense ECM elasticity. RHO family GTPases are significant regulators of the actin cytoskeleton and of mechanotransduction. It has been found that RHO-mediated contractility and independent actomyosin contractility regulates osteogenic differentiation.68, 152 Increased Rac1 activity was also reported to enhance smooth muscle cell differentiation.153 YAP/TAZ has been considered as key mediators in cell responses to substrate stiffness, such as that in viscoelastic regulation of cell behaviours. For example, stress relaxation of a substrate has been found to lead to increased nuclear translocation of YAP, resulting in increased U2OS cell proliferation.74 Further, the cell spreading, proliferation, and osteogenic differentiation of MSCs was enhanced on gels with faster stress relaxation, which was proven to be mediated through integrin-based adhesion, actomyosin contractility, and nuclear localization of YAP.68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume. ...
... 68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume. ...
Positive alterations of viscoelastic and geometric properties in ovariectomized rat femurs with concurrent administration of ibandronate and PTH
1
2013
... Stiffness or elasticity has been recognized as the most important parameter in evaluating the mechanical properties of biological tissues.8, 18, 19 Yet, it should be noted that the mechanical properties of tissues or organs are dynamic and constantly changing showing spatio-temporal characteristics. Viscoelasticity is an important mechanical parameter with a time dimension. As a common and basic mechanical feature of native tissues, viscoelasticity has been widely examined in living tissues and organs, including skin, muscle, cartilage, blood vessels, tendons, brain, liver, breast, and embryo.9 The viscoelasticity of tissues is mainly the product of the ECM and resident cells. ECM is rich in biological macromolecular substances, such as proteoglycan, hyaluronic acid, collagen, fibrin and laminin, whose mechanical behaviours show time-dependent characteristics. The diversity in biochemical components and tissue-specific structures contributes to the characteristic ECM stress relaxation, i.e., viscoelasticity. The viscoelasticity of a tissue plays an important role in the normal physiological function of the organism, especially in the motor system to protect tissues and cells under persistent or irregular high stress. For example, the viscoelastic connections of bone, muscle and connective tissue cushion stress conduction and increase energy storage and dissipation.65 In addition, changes in the viscoelasticity of healthy tissues are also considered relevant to the occurrence and development of pathological processes, including liver fibrosis, progression of breast cancer, scleroderma, and osteoporotic deterioration.66-69 Consequently, thorough examination of the viscoelasticity of living tissues would lead us to fully understand the mechanism of physiological activities and pathological disorders, so as to guide the design of tissue-mimicking viscoelastic scaffolds for tissue regeneration. In this article, we discuss the underlying principles and recent advances in the viscoelasticity-dependent regulation of various cell behaviours, including cell adhesion, diffusion, migration, proliferation, and stem-cell differentiation. ...
The influence of substrate creep on mesenchymal stem cell behaviour and phenotype
3
2011
... Polyacrylamide hydrogels, with varying loss moduli caused by movement of the loose ends of polymer chains, was an early approach to explore the effect of viscoelasticity on hMSC behaviour. The results highlighted that substrate creep was an important mechanical property of a biomaterial system to encourage cell growth and differentiation.70 Recently, Charrier et al.71 reported a type of soft, viscoelastic polyacrylamide hydrogel with independently-tuneable elasticity and viscous dissipation, due to the movements of entangled linear polyacrylamide which was sterically entrapped in the polyacrylamide crosslinked networks. They demonstrated that viscous dissipation is a determinant of cell phenotype and tissue homeostasis.71 ...
... Recent studies showed that cell adhesion can be influenced by the viscoelasticity of the substrate as well its elasticity. For example, when U2OS cells were cultured on stress-relaxed alginate hydrogels, greater cellular FA formation was observed compared with that on purely elastic substrates, as indicated by the punctate localization of paxillin.72 Subsequently, the same group studied cell adhesion of MSCs in RGD-coupled alginate hydrogels with various rates of stress relaxation. They found that localization of integrin-β1 was increased at the periphery of the cell in faster relaxation gels.73 However, at any level of stress relaxation, there was no localisation of paxillin to the periphery of the cells, indicating that no conventional FAs were formed in this 3D viscoelastic hydrogel. In another study, the same group developed a novel viscoelastic matrix composed of hyaluronic acid and collagen type I,81 and found that FAs were promoted with higher expression of integrin and paxillin, while they were minimally-expressed and diffuse in the corresponding elastic substrate, due to the synergistic effect of viscoelasticity and fibrillar architecture. However, there have been some contradictory findings in other viscoelastic systems. In 2019, an alginate-PEG hydrogel with tuneable stress relaxation was developed for 3D culture of MSCs.74 It was found that localisation of integrin-β1 increased with faster relaxation, exhibiting higher levels of phosphorylated FA kinase (FAK) and paxillin. On polyacrylamide-based hydrogels with similar initial storage moduli and various loss moduli, paxillin clusters were much smaller on viscoelastic gels than on elastic gels, suggesting that energy dissipation of the substrate inhibits formation of FAs.70 ...
... When hMSCs were cultured on a polyacrylamide gel system with varying loss modulus, the multilineage differentiation of hMSCs was enhanced in specific inductive media on high-loss modulus substrates. These results highlighted that substrate loss modulus was a significant mechanical parameter to be tailored for biomaterials in tissue engineering.70 Ionically-crosslinked alginate hydrogels are a typical viscoelastic hydrogel. A previous study found that MSCs undergo major adipogenic differentiation when the initial modulus is 1–10 kPa, and osteogenic differentiation when the initial modulus is 11–30 kPa. Chaudhuri et al.73 developed viscoelastic alginate hydrogels with various stress relaxation properties and initial elastic moduli, to study the differentiation of encapsulated MSCs. They found that MSCs exhibited primarily adipogenic differentiation at all timescales of stress relaxation when the initial elastic modulus was ~9 kPa, while the level of adipogenesis was decreased in rapidly-relaxing gels. In contrast, no adipogenic differentiation was observed when the initial elastic modulus was ~17 kPa, and osteogenic differentiation was significantly enhanced in faster stress relaxing gels. In addition, an interconnected bone-like matrix was formed in rapidly-relaxing gels with relaxation time at ~1 minute, indicating that faster stress relaxation promoted both osteogenesis and bone-forming activity. A subsequent study confirmed that faster relaxation also led to enhanced osteogenic differentiation of MSCs on PEG covalently-grafted alginate-PEG hydrogels.74 ...
Control of cell morphology and differentiation by substrates with independently tunable elasticity and viscous dissipation
3
2018
... Polyacrylamide hydrogels, with varying loss moduli caused by movement of the loose ends of polymer chains, was an early approach to explore the effect of viscoelasticity on hMSC behaviour. The results highlighted that substrate creep was an important mechanical property of a biomaterial system to encourage cell growth and differentiation.70 Recently, Charrier et al.71 reported a type of soft, viscoelastic polyacrylamide hydrogel with independently-tuneable elasticity and viscous dissipation, due to the movements of entangled linear polyacrylamide which was sterically entrapped in the polyacrylamide crosslinked networks. They demonstrated that viscous dissipation is a determinant of cell phenotype and tissue homeostasis.71 ...
... 71 ...
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
... However, most viscoelastic adjustable hydrogels were developed by creating networks with specific crosslinkers including physically non-covalent crosslinkers (e.g., ionic interaction,72-75 hydrogen bonding,76 guest-host interactions,77 etc.) and chemically dynamic covalent crosslinkers (e.g., acetyl exchange, disulphide bonds, hydrazone/oxime exchange, Diels-Alder reaction, etc.)75, 78-85 (Additional Table 2). ...
... Recent studies showed that cell adhesion can be influenced by the viscoelasticity of the substrate as well its elasticity. For example, when U2OS cells were cultured on stress-relaxed alginate hydrogels, greater cellular FA formation was observed compared with that on purely elastic substrates, as indicated by the punctate localization of paxillin.72 Subsequently, the same group studied cell adhesion of MSCs in RGD-coupled alginate hydrogels with various rates of stress relaxation. They found that localization of integrin-β1 was increased at the periphery of the cell in faster relaxation gels.73 However, at any level of stress relaxation, there was no localisation of paxillin to the periphery of the cells, indicating that no conventional FAs were formed in this 3D viscoelastic hydrogel. In another study, the same group developed a novel viscoelastic matrix composed of hyaluronic acid and collagen type I,81 and found that FAs were promoted with higher expression of integrin and paxillin, while they were minimally-expressed and diffuse in the corresponding elastic substrate, due to the synergistic effect of viscoelasticity and fibrillar architecture. However, there have been some contradictory findings in other viscoelastic systems. In 2019, an alginate-PEG hydrogel with tuneable stress relaxation was developed for 3D culture of MSCs.74 It was found that localisation of integrin-β1 increased with faster relaxation, exhibiting higher levels of phosphorylated FA kinase (FAK) and paxillin. On polyacrylamide-based hydrogels with similar initial storage moduli and various loss moduli, paxillin clusters were much smaller on viscoelastic gels than on elastic gels, suggesting that energy dissipation of the substrate inhibits formation of FAs.70 ...
... When a cell attaches to a substrate, it receives signals via integrins, after which actin polymerises to cause movement and extension of the cell membrane, while myosin contraction and membrane tension forces the cell to maintain tensional homeostasis, determining the cell spread area. A number of efforts have been devoted to investigating the effect of viscoelasticity on cell spreading in both two-dimensional (2D) and 3D culture. On 2D viscoelastic substrates, cell spreading was increased compared with elastic substrates, e.g. MSCs on collagen type I-coated polyacrylamide hydrogels, or U2OS cells72 and myoblasts83 on Arg-Gly-Asp (RGD)-modified alginate hydrogels. Conversely, when NIH-3T3 fibroblasts were cultured on boronate ester-based viscoelastic hydrogels, in all of the cell area and nuclear area, FA was lower than that on elastic hydrogels with the same storage modulus. In addition, with the change of stress relaxation time, the morphology of the cells will also change. Mostly, cells spread better and develop a less-rounded shaped on a fast stress relaxing substrate than on a slow stress relaxed substrate. For example, on an RGD-modified viscoelastic alginate hydrogel, fibroblasts and BMSCs showed greater cell expansion when τ1/2 was reduced from 1 hour to 1 minute.73 In 3D viscoelastic substrates, a similar phenomenon was observed. For example, in an RGD-coupled alginate-PEG hydrogel, the spreading of 3T3 fibroblasts was inhibited and cells with a spherical morphology were observed in slow stress relaxed gels, while cells spread more in the fast stress relaxing gels.73 ...
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Hydrogels with tunable stress relaxation regulate stem cell fate and activity
7
2016
... Alginate, a natural material, has been used to fabricate viscoelastic matrices for cell culture. The viscoelasticity can be controlled by changing the ratio of ionic cross-linker to covalent cross-linker, whose stress relaxation occurrs as a result of crosslinker unbinding.86 Based on this, Chaudhuri’s group.73, 87 reported that alginate hydrogels with various stress relaxation properties were obtained by changing the molecular weight of alginate or covalent incorporation of short PEG chains, which modulated the density of ionic crosslinks or led to a looser network in gels. Most recently, the same group found that stress relaxation of alginate hydrogel was simply and precisely modulated by changing the molecular weight and concentration of PEG.74 Increasing PEG concentration and molecular weight led to faster stress relaxation, higher loss modulus and increased creep. In order to simulate the fibrillar architecture of natural ECM, an interpenetrating network of click-functionalised alginate and fibrillar collagen type I was fabricated as a viscoelastic artificial ECM.75 Timing sequential ionic and covalent crosslinking allowed collagen type I to self-assemble into fibrous structures within the network. The stiffness and viscoelasticity was tuned by changing the mode and magnitude of crosslinking, without changing the micro-scale architecture of the hydrogels. The above viscoelastic alginate hydrogels were mostly ionically cross-linked, mainly using physical approaches to regulate viscoelasticity. Besides the ionic interaction, other physically cross-linked hydrogels have also been explored to form viscoelastic hydrogels including hydrogen bonding and guest-host interaction. ...
... Recent studies showed that cell adhesion can be influenced by the viscoelasticity of the substrate as well its elasticity. For example, when U2OS cells were cultured on stress-relaxed alginate hydrogels, greater cellular FA formation was observed compared with that on purely elastic substrates, as indicated by the punctate localization of paxillin.72 Subsequently, the same group studied cell adhesion of MSCs in RGD-coupled alginate hydrogels with various rates of stress relaxation. They found that localization of integrin-β1 was increased at the periphery of the cell in faster relaxation gels.73 However, at any level of stress relaxation, there was no localisation of paxillin to the periphery of the cells, indicating that no conventional FAs were formed in this 3D viscoelastic hydrogel. In another study, the same group developed a novel viscoelastic matrix composed of hyaluronic acid and collagen type I,81 and found that FAs were promoted with higher expression of integrin and paxillin, while they were minimally-expressed and diffuse in the corresponding elastic substrate, due to the synergistic effect of viscoelasticity and fibrillar architecture. However, there have been some contradictory findings in other viscoelastic systems. In 2019, an alginate-PEG hydrogel with tuneable stress relaxation was developed for 3D culture of MSCs.74 It was found that localisation of integrin-β1 increased with faster relaxation, exhibiting higher levels of phosphorylated FA kinase (FAK) and paxillin. On polyacrylamide-based hydrogels with similar initial storage moduli and various loss moduli, paxillin clusters were much smaller on viscoelastic gels than on elastic gels, suggesting that energy dissipation of the substrate inhibits formation of FAs.70 ...
... When a cell attaches to a substrate, it receives signals via integrins, after which actin polymerises to cause movement and extension of the cell membrane, while myosin contraction and membrane tension forces the cell to maintain tensional homeostasis, determining the cell spread area. A number of efforts have been devoted to investigating the effect of viscoelasticity on cell spreading in both two-dimensional (2D) and 3D culture. On 2D viscoelastic substrates, cell spreading was increased compared with elastic substrates, e.g. MSCs on collagen type I-coated polyacrylamide hydrogels, or U2OS cells72 and myoblasts83 on Arg-Gly-Asp (RGD)-modified alginate hydrogels. Conversely, when NIH-3T3 fibroblasts were cultured on boronate ester-based viscoelastic hydrogels, in all of the cell area and nuclear area, FA was lower than that on elastic hydrogels with the same storage modulus. In addition, with the change of stress relaxation time, the morphology of the cells will also change. Mostly, cells spread better and develop a less-rounded shaped on a fast stress relaxing substrate than on a slow stress relaxed substrate. For example, on an RGD-modified viscoelastic alginate hydrogel, fibroblasts and BMSCs showed greater cell expansion when τ1/2 was reduced from 1 hour to 1 minute.73 In 3D viscoelastic substrates, a similar phenomenon was observed. For example, in an RGD-coupled alginate-PEG hydrogel, the spreading of 3T3 fibroblasts was inhibited and cells with a spherical morphology were observed in slow stress relaxed gels, while cells spread more in the fast stress relaxing gels.73 ...
... 73 ...
... The proliferation of various resident cells in a viscoelastic microenvironment was also found to be enhanced and regulated. Several studies demonstrated that hydrogel viscoelasticity has an obvious effect in promoting spreading and proliferation of fibroblasts in 2D or 3D culture.73, 88, 89 Similarly, when hMSCs were cultured on polyacrylamide-based viscoelastic hydrogels, the proliferation rate of hMSCs was significantly promoted by faster stress relaxation times.64 On an oxime cross-linked alginate hydrogel,82 2PK3 cells showed the same trend, with cells growing better in hydrogels with fast stress relaxation. All results indicate that stress relaxation time-dependent regulation may have beneficial effects on proliferation. ...
... When hMSCs were cultured on a polyacrylamide gel system with varying loss modulus, the multilineage differentiation of hMSCs was enhanced in specific inductive media on high-loss modulus substrates. These results highlighted that substrate loss modulus was a significant mechanical parameter to be tailored for biomaterials in tissue engineering.70 Ionically-crosslinked alginate hydrogels are a typical viscoelastic hydrogel. A previous study found that MSCs undergo major adipogenic differentiation when the initial modulus is 1–10 kPa, and osteogenic differentiation when the initial modulus is 11–30 kPa. Chaudhuri et al.73 developed viscoelastic alginate hydrogels with various stress relaxation properties and initial elastic moduli, to study the differentiation of encapsulated MSCs. They found that MSCs exhibited primarily adipogenic differentiation at all timescales of stress relaxation when the initial elastic modulus was ~9 kPa, while the level of adipogenesis was decreased in rapidly-relaxing gels. In contrast, no adipogenic differentiation was observed when the initial elastic modulus was ~17 kPa, and osteogenic differentiation was significantly enhanced in faster stress relaxing gels. In addition, an interconnected bone-like matrix was formed in rapidly-relaxing gels with relaxation time at ~1 minute, indicating that faster stress relaxation promoted both osteogenesis and bone-forming activity. A subsequent study confirmed that faster relaxation also led to enhanced osteogenic differentiation of MSCs on PEG covalently-grafted alginate-PEG hydrogels.74 ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Varying PEG density to control stress relaxation in alginate-PEG hydrogels for 3D cell culture studies
5
2019
... Alginate, a natural material, has been used to fabricate viscoelastic matrices for cell culture. The viscoelasticity can be controlled by changing the ratio of ionic cross-linker to covalent cross-linker, whose stress relaxation occurrs as a result of crosslinker unbinding.86 Based on this, Chaudhuri’s group.73, 87 reported that alginate hydrogels with various stress relaxation properties were obtained by changing the molecular weight of alginate or covalent incorporation of short PEG chains, which modulated the density of ionic crosslinks or led to a looser network in gels. Most recently, the same group found that stress relaxation of alginate hydrogel was simply and precisely modulated by changing the molecular weight and concentration of PEG.74 Increasing PEG concentration and molecular weight led to faster stress relaxation, higher loss modulus and increased creep. In order to simulate the fibrillar architecture of natural ECM, an interpenetrating network of click-functionalised alginate and fibrillar collagen type I was fabricated as a viscoelastic artificial ECM.75 Timing sequential ionic and covalent crosslinking allowed collagen type I to self-assemble into fibrous structures within the network. The stiffness and viscoelasticity was tuned by changing the mode and magnitude of crosslinking, without changing the micro-scale architecture of the hydrogels. The above viscoelastic alginate hydrogels were mostly ionically cross-linked, mainly using physical approaches to regulate viscoelasticity. Besides the ionic interaction, other physically cross-linked hydrogels have also been explored to form viscoelastic hydrogels including hydrogen bonding and guest-host interaction. ...
... Recent studies showed that cell adhesion can be influenced by the viscoelasticity of the substrate as well its elasticity. For example, when U2OS cells were cultured on stress-relaxed alginate hydrogels, greater cellular FA formation was observed compared with that on purely elastic substrates, as indicated by the punctate localization of paxillin.72 Subsequently, the same group studied cell adhesion of MSCs in RGD-coupled alginate hydrogels with various rates of stress relaxation. They found that localization of integrin-β1 was increased at the periphery of the cell in faster relaxation gels.73 However, at any level of stress relaxation, there was no localisation of paxillin to the periphery of the cells, indicating that no conventional FAs were formed in this 3D viscoelastic hydrogel. In another study, the same group developed a novel viscoelastic matrix composed of hyaluronic acid and collagen type I,81 and found that FAs were promoted with higher expression of integrin and paxillin, while they were minimally-expressed and diffuse in the corresponding elastic substrate, due to the synergistic effect of viscoelasticity and fibrillar architecture. However, there have been some contradictory findings in other viscoelastic systems. In 2019, an alginate-PEG hydrogel with tuneable stress relaxation was developed for 3D culture of MSCs.74 It was found that localisation of integrin-β1 increased with faster relaxation, exhibiting higher levels of phosphorylated FA kinase (FAK) and paxillin. On polyacrylamide-based hydrogels with similar initial storage moduli and various loss moduli, paxillin clusters were much smaller on viscoelastic gels than on elastic gels, suggesting that energy dissipation of the substrate inhibits formation of FAs.70 ...
... When hMSCs were cultured on a polyacrylamide gel system with varying loss modulus, the multilineage differentiation of hMSCs was enhanced in specific inductive media on high-loss modulus substrates. These results highlighted that substrate loss modulus was a significant mechanical parameter to be tailored for biomaterials in tissue engineering.70 Ionically-crosslinked alginate hydrogels are a typical viscoelastic hydrogel. A previous study found that MSCs undergo major adipogenic differentiation when the initial modulus is 1–10 kPa, and osteogenic differentiation when the initial modulus is 11–30 kPa. Chaudhuri et al.73 developed viscoelastic alginate hydrogels with various stress relaxation properties and initial elastic moduli, to study the differentiation of encapsulated MSCs. They found that MSCs exhibited primarily adipogenic differentiation at all timescales of stress relaxation when the initial elastic modulus was ~9 kPa, while the level of adipogenesis was decreased in rapidly-relaxing gels. In contrast, no adipogenic differentiation was observed when the initial elastic modulus was ~17 kPa, and osteogenic differentiation was significantly enhanced in faster stress relaxing gels. In addition, an interconnected bone-like matrix was formed in rapidly-relaxing gels with relaxation time at ~1 minute, indicating that faster stress relaxation promoted both osteogenesis and bone-forming activity. A subsequent study confirmed that faster relaxation also led to enhanced osteogenic differentiation of MSCs on PEG covalently-grafted alginate-PEG hydrogels.74 ...
... The methods used to explore the molecular mechanism of how cells sense ECM viscoelasticity are similar to those used in studying how cells sense ECM elasticity. RHO family GTPases are significant regulators of the actin cytoskeleton and of mechanotransduction. It has been found that RHO-mediated contractility and independent actomyosin contractility regulates osteogenic differentiation.68, 152 Increased Rac1 activity was also reported to enhance smooth muscle cell differentiation.153 YAP/TAZ has been considered as key mediators in cell responses to substrate stiffness, such as that in viscoelastic regulation of cell behaviours. For example, stress relaxation of a substrate has been found to lead to increased nuclear translocation of YAP, resulting in increased U2OS cell proliferation.74 Further, the cell spreading, proliferation, and osteogenic differentiation of MSCs was enhanced on gels with faster stress relaxation, which was proven to be mediated through integrin-based adhesion, actomyosin contractility, and nuclear localization of YAP.68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume. ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Sequential modes of crosslinking tune viscoelasticity of cell-instructive hydrogels
5
2019
... However, most viscoelastic adjustable hydrogels were developed by creating networks with specific crosslinkers including physically non-covalent crosslinkers (e.g., ionic interaction,72-75 hydrogen bonding,76 guest-host interactions,77 etc.) and chemically dynamic covalent crosslinkers (e.g., acetyl exchange, disulphide bonds, hydrazone/oxime exchange, Diels-Alder reaction, etc.)75, 78-85 (Additional Table 2). ...
... 75, 78-85 (Additional Table 2). ...
... Alginate, a natural material, has been used to fabricate viscoelastic matrices for cell culture. The viscoelasticity can be controlled by changing the ratio of ionic cross-linker to covalent cross-linker, whose stress relaxation occurrs as a result of crosslinker unbinding.86 Based on this, Chaudhuri’s group.73, 87 reported that alginate hydrogels with various stress relaxation properties were obtained by changing the molecular weight of alginate or covalent incorporation of short PEG chains, which modulated the density of ionic crosslinks or led to a looser network in gels. Most recently, the same group found that stress relaxation of alginate hydrogel was simply and precisely modulated by changing the molecular weight and concentration of PEG.74 Increasing PEG concentration and molecular weight led to faster stress relaxation, higher loss modulus and increased creep. In order to simulate the fibrillar architecture of natural ECM, an interpenetrating network of click-functionalised alginate and fibrillar collagen type I was fabricated as a viscoelastic artificial ECM.75 Timing sequential ionic and covalent crosslinking allowed collagen type I to self-assemble into fibrous structures within the network. The stiffness and viscoelasticity was tuned by changing the mode and magnitude of crosslinking, without changing the micro-scale architecture of the hydrogels. The above viscoelastic alginate hydrogels were mostly ionically cross-linked, mainly using physical approaches to regulate viscoelasticity. Besides the ionic interaction, other physically cross-linked hydrogels have also been explored to form viscoelastic hydrogels including hydrogen bonding and guest-host interaction. ...
... The methods used to explore the molecular mechanism of how cells sense ECM viscoelasticity are similar to those used in studying how cells sense ECM elasticity. RHO family GTPases are significant regulators of the actin cytoskeleton and of mechanotransduction. It has been found that RHO-mediated contractility and independent actomyosin contractility regulates osteogenic differentiation.68, 152 Increased Rac1 activity was also reported to enhance smooth muscle cell differentiation.153 YAP/TAZ has been considered as key mediators in cell responses to substrate stiffness, such as that in viscoelastic regulation of cell behaviours. For example, stress relaxation of a substrate has been found to lead to increased nuclear translocation of YAP, resulting in increased U2OS cell proliferation.74 Further, the cell spreading, proliferation, and osteogenic differentiation of MSCs was enhanced on gels with faster stress relaxation, which was proven to be mediated through integrin-based adhesion, actomyosin contractility, and nuclear localization of YAP.68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume. ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Strain-enhanced stress relaxation impacts nonlinear elasticity in collagen gels
2
2016
... However, most viscoelastic adjustable hydrogels were developed by creating networks with specific crosslinkers including physically non-covalent crosslinkers (e.g., ionic interaction,72-75 hydrogen bonding,76 guest-host interactions,77 etc.) and chemically dynamic covalent crosslinkers (e.g., acetyl exchange, disulphide bonds, hydrazone/oxime exchange, Diels-Alder reaction, etc.)75, 78-85 (Additional Table 2). ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Reversible control of network properties in azobenzene-containing hyaluronic acid-based hydrogels
2
2018
... However, most viscoelastic adjustable hydrogels were developed by creating networks with specific crosslinkers including physically non-covalent crosslinkers (e.g., ionic interaction,72-75 hydrogen bonding,76 guest-host interactions,77 etc.) and chemically dynamic covalent crosslinkers (e.g., acetyl exchange, disulphide bonds, hydrazone/oxime exchange, Diels-Alder reaction, etc.)75, 78-85 (Additional Table 2). ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Adaptable boronate ester hydrogels with tunable viscoelastic spectra to probe timescale dependent mechanotransduction
2
2019
... However, most viscoelastic adjustable hydrogels were developed by creating networks with specific crosslinkers including physically non-covalent crosslinkers (e.g., ionic interaction,72-75 hydrogen bonding,76 guest-host interactions,77 etc.) and chemically dynamic covalent crosslinkers (e.g., acetyl exchange, disulphide bonds, hydrazone/oxime exchange, Diels-Alder reaction, etc.)75, 78-85 (Additional Table 2). ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Adaptable fast relaxing boronate-based hydrogels for probing cell-matrix interactions
1
2018
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Spatiotemporal control of viscoelasticity in phototunable hyaluronic acid hydrogels
2
2019
... Although many strategies have been developed to tune the viscoelasticity of hydrogels as mentioned above, there is still a huge demand to create biomaterials with tuneable viscoelasticity to mimic the features of native tissues. Recently, in order to simulate the dynamic and heterogenous nature of tissues, Hui et al.80 developed a spatiotemporal controllable viscoelastic hydrogel based on phototunable hyaluronic acid, which showed great promise in simulating the range of viscoelasticities representing healthy and diseased cellular environments. ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Stress relaxing hyaluronic acid-collagen hydrogels promote cell spreading, fiber remodeling, and focal adhesion formation in 3D cell culture
2
2018
... Recent studies showed that cell adhesion can be influenced by the viscoelasticity of the substrate as well its elasticity. For example, when U2OS cells were cultured on stress-relaxed alginate hydrogels, greater cellular FA formation was observed compared with that on purely elastic substrates, as indicated by the punctate localization of paxillin.72 Subsequently, the same group studied cell adhesion of MSCs in RGD-coupled alginate hydrogels with various rates of stress relaxation. They found that localization of integrin-β1 was increased at the periphery of the cell in faster relaxation gels.73 However, at any level of stress relaxation, there was no localisation of paxillin to the periphery of the cells, indicating that no conventional FAs were formed in this 3D viscoelastic hydrogel. In another study, the same group developed a novel viscoelastic matrix composed of hyaluronic acid and collagen type I,81 and found that FAs were promoted with higher expression of integrin and paxillin, while they were minimally-expressed and diffuse in the corresponding elastic substrate, due to the synergistic effect of viscoelasticity and fibrillar architecture. However, there have been some contradictory findings in other viscoelastic systems. In 2019, an alginate-PEG hydrogel with tuneable stress relaxation was developed for 3D culture of MSCs.74 It was found that localisation of integrin-β1 increased with faster relaxation, exhibiting higher levels of phosphorylated FA kinase (FAK) and paxillin. On polyacrylamide-based hydrogels with similar initial storage moduli and various loss moduli, paxillin clusters were much smaller on viscoelastic gels than on elastic gels, suggesting that energy dissipation of the substrate inhibits formation of FAs.70 ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Oxime cross-linked alginate hydrogels with tunable stress relaxation
4
2019
... In addition, dynamically-reversible cross-links in hydrogels can undergo rapid breakage and re-formation when subjected to stress. The viscoelasticity of hydrogels can be adjusted by varying the density of dynamic chemical cross-linkers or adjusting the exchange rate of reversible cross-linkers. Imine and hydrazone bonds are reversible bonds which have been successfully used to fabricate many viscoelastic hydrogels, including hyaluronic acid, PEG, and alginate. For example, McKinnon et al.83 developed PEG-based viscoelastic hydrogels based on reversible hydrazone bonds with two different affinities. The stress relaxation rate (τ1/2) was tuned in the range of ~5–33,600 seconds, by changing the arm number of PEG or the ratio of aliphatic aldehyde to benzaldehyde crosslinkers. Further, the same group prepared 8-arm PEG viscoelastic hydrogels with stress relaxation times in the range of three orders of magnitude to check the effect on chondrocyte functions for cartilage tissue engineering. Recently, Sánchez-Morán et al.82 fabricated novel alginate hydrogels with tuneable stress relaxation based on reversible oxime bonds. They first prepared alkoxy-amine functionalized alginate, and then mixed this with aldehyde-containing alginate to form oxime cross-linked hydrogels. The stress relaxation and mechanical properties were adjusted by changing the concentration, mixing ratios, degree of oxidation of aldehyde-containing oxidized alginate or environmental factors (e.g. pH, temperature, and use of catalyst). Most interestingly, this study provided novel methods of preparing viscoelastic hydrogels in microbeads or microthread geometries.82 There are also other reversible covalent crosslinking methods, e.g. thioester, boronate ester, or Diels-Alder reaction, which can be used to fabricate dynamic hydrogels. For example, Brown et al.84 presented a viscoelastic hydrogel based on thioester exchange. The τ1/2 was tuneable across several orders of magnitude by controlling gel stoichiometry, pH, and crosslinker structure. ...
... 82 There are also other reversible covalent crosslinking methods, e.g. thioester, boronate ester, or Diels-Alder reaction, which can be used to fabricate dynamic hydrogels. For example, Brown et al.84 presented a viscoelastic hydrogel based on thioester exchange. The τ1/2 was tuneable across several orders of magnitude by controlling gel stoichiometry, pH, and crosslinker structure. ...
... The proliferation of various resident cells in a viscoelastic microenvironment was also found to be enhanced and regulated. Several studies demonstrated that hydrogel viscoelasticity has an obvious effect in promoting spreading and proliferation of fibroblasts in 2D or 3D culture.73, 88, 89 Similarly, when hMSCs were cultured on polyacrylamide-based viscoelastic hydrogels, the proliferation rate of hMSCs was significantly promoted by faster stress relaxation times.64 On an oxime cross-linked alginate hydrogel,82 2PK3 cells showed the same trend, with cells growing better in hydrogels with fast stress relaxation. All results indicate that stress relaxation time-dependent regulation may have beneficial effects on proliferation. ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Biophysically defined and cytocompatible covalently adaptable networks as viscoelastic 3D cell culture systems
4
2014
... In addition, dynamically-reversible cross-links in hydrogels can undergo rapid breakage and re-formation when subjected to stress. The viscoelasticity of hydrogels can be adjusted by varying the density of dynamic chemical cross-linkers or adjusting the exchange rate of reversible cross-linkers. Imine and hydrazone bonds are reversible bonds which have been successfully used to fabricate many viscoelastic hydrogels, including hyaluronic acid, PEG, and alginate. For example, McKinnon et al.83 developed PEG-based viscoelastic hydrogels based on reversible hydrazone bonds with two different affinities. The stress relaxation rate (τ1/2) was tuned in the range of ~5–33,600 seconds, by changing the arm number of PEG or the ratio of aliphatic aldehyde to benzaldehyde crosslinkers. Further, the same group prepared 8-arm PEG viscoelastic hydrogels with stress relaxation times in the range of three orders of magnitude to check the effect on chondrocyte functions for cartilage tissue engineering. Recently, Sánchez-Morán et al.82 fabricated novel alginate hydrogels with tuneable stress relaxation based on reversible oxime bonds. They first prepared alkoxy-amine functionalized alginate, and then mixed this with aldehyde-containing alginate to form oxime cross-linked hydrogels. The stress relaxation and mechanical properties were adjusted by changing the concentration, mixing ratios, degree of oxidation of aldehyde-containing oxidized alginate or environmental factors (e.g. pH, temperature, and use of catalyst). Most interestingly, this study provided novel methods of preparing viscoelastic hydrogels in microbeads or microthread geometries.82 There are also other reversible covalent crosslinking methods, e.g. thioester, boronate ester, or Diels-Alder reaction, which can be used to fabricate dynamic hydrogels. For example, Brown et al.84 presented a viscoelastic hydrogel based on thioester exchange. The τ1/2 was tuneable across several orders of magnitude by controlling gel stoichiometry, pH, and crosslinker structure. ...
... When a cell attaches to a substrate, it receives signals via integrins, after which actin polymerises to cause movement and extension of the cell membrane, while myosin contraction and membrane tension forces the cell to maintain tensional homeostasis, determining the cell spread area. A number of efforts have been devoted to investigating the effect of viscoelasticity on cell spreading in both two-dimensional (2D) and 3D culture. On 2D viscoelastic substrates, cell spreading was increased compared with elastic substrates, e.g. MSCs on collagen type I-coated polyacrylamide hydrogels, or U2OS cells72 and myoblasts83 on Arg-Gly-Asp (RGD)-modified alginate hydrogels. Conversely, when NIH-3T3 fibroblasts were cultured on boronate ester-based viscoelastic hydrogels, in all of the cell area and nuclear area, FA was lower than that on elastic hydrogels with the same storage modulus. In addition, with the change of stress relaxation time, the morphology of the cells will also change. Mostly, cells spread better and develop a less-rounded shaped on a fast stress relaxing substrate than on a slow stress relaxed substrate. For example, on an RGD-modified viscoelastic alginate hydrogel, fibroblasts and BMSCs showed greater cell expansion when τ1/2 was reduced from 1 hour to 1 minute.73 In 3D viscoelastic substrates, a similar phenomenon was observed. For example, in an RGD-coupled alginate-PEG hydrogel, the spreading of 3T3 fibroblasts was inhibited and cells with a spherical morphology were observed in slow stress relaxed gels, while cells spread more in the fast stress relaxing gels.73 ...
... The methods used to explore the molecular mechanism of how cells sense ECM viscoelasticity are similar to those used in studying how cells sense ECM elasticity. RHO family GTPases are significant regulators of the actin cytoskeleton and of mechanotransduction. It has been found that RHO-mediated contractility and independent actomyosin contractility regulates osteogenic differentiation.68, 152 Increased Rac1 activity was also reported to enhance smooth muscle cell differentiation.153 YAP/TAZ has been considered as key mediators in cell responses to substrate stiffness, such as that in viscoelastic regulation of cell behaviours. For example, stress relaxation of a substrate has been found to lead to increased nuclear translocation of YAP, resulting in increased U2OS cell proliferation.74 Further, the cell spreading, proliferation, and osteogenic differentiation of MSCs was enhanced on gels with faster stress relaxation, which was proven to be mediated through integrin-based adhesion, actomyosin contractility, and nuclear localization of YAP.68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume. ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Photopolymerized dynamic hydrogels with tunable viscoelastic properties through thioester exchange
2
2018
... In addition, dynamically-reversible cross-links in hydrogels can undergo rapid breakage and re-formation when subjected to stress. The viscoelasticity of hydrogels can be adjusted by varying the density of dynamic chemical cross-linkers or adjusting the exchange rate of reversible cross-linkers. Imine and hydrazone bonds are reversible bonds which have been successfully used to fabricate many viscoelastic hydrogels, including hyaluronic acid, PEG, and alginate. For example, McKinnon et al.83 developed PEG-based viscoelastic hydrogels based on reversible hydrazone bonds with two different affinities. The stress relaxation rate (τ1/2) was tuned in the range of ~5–33,600 seconds, by changing the arm number of PEG or the ratio of aliphatic aldehyde to benzaldehyde crosslinkers. Further, the same group prepared 8-arm PEG viscoelastic hydrogels with stress relaxation times in the range of three orders of magnitude to check the effect on chondrocyte functions for cartilage tissue engineering. Recently, Sánchez-Morán et al.82 fabricated novel alginate hydrogels with tuneable stress relaxation based on reversible oxime bonds. They first prepared alkoxy-amine functionalized alginate, and then mixed this with aldehyde-containing alginate to form oxime cross-linked hydrogels. The stress relaxation and mechanical properties were adjusted by changing the concentration, mixing ratios, degree of oxidation of aldehyde-containing oxidized alginate or environmental factors (e.g. pH, temperature, and use of catalyst). Most interestingly, this study provided novel methods of preparing viscoelastic hydrogels in microbeads or microthread geometries.82 There are also other reversible covalent crosslinking methods, e.g. thioester, boronate ester, or Diels-Alder reaction, which can be used to fabricate dynamic hydrogels. For example, Brown et al.84 presented a viscoelastic hydrogel based on thioester exchange. The τ1/2 was tuneable across several orders of magnitude by controlling gel stoichiometry, pH, and crosslinker structure. ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Tailoring supramolecular guest-host hydrogel viscoelasticity with covalent fibrinogen double networks
2
2019
... However, most viscoelastic adjustable hydrogels were developed by creating networks with specific crosslinkers including physically non-covalent crosslinkers (e.g., ionic interaction,72-75 hydrogen bonding,76 guest-host interactions,77 etc.) and chemically dynamic covalent crosslinkers (e.g., acetyl exchange, disulphide bonds, hydrazone/oxime exchange, Diels-Alder reaction, etc.)75, 78-85 (Additional Table 2). ...
... Scaffolds with tuneable viscoelasticity through various crosslinkers and their effects on cells.
Biomaterials
Fabrication method
Viscoelasticity
Effects on cell behaviours
Cell source
Reference
Alginate hydrogels
Prepared by ionic crosslinking of alginate
Obtained by covalently or ionically crosslinking alginate gels with the same initial Young’s modulus by adjusting the concentration of crosslinker
Both computational modelling and experimental studies revealed that spreading of cells cultured on soft substrates that exhibit stress relaxation is greater than cell spreading on elastic substrates of the same modulus, but similar to that of cells spreading on stiffer elastic substrates
U2OS & 3T3 fibroblasts
72
Prepared by ionic crosslinking of alginate
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~1 minute to ~1 hour by controlling the molecular weight of alginate
Cell spreading, proliferation, and osteogenic differentiation of MSCs were all enhanced in cells cultured in gels with faster relaxation
MSCs
73
Alginate-PEG hydrogels
Prepared by ionic crosslinking of PEG-functionalised alginate
PEG acts as a spacer to provide a steric spacing of crosslinking zones in alginate. Increased concentration and molecular weight of the PEG resulted in faster stress relaxation, a high loss modulus, and increased creep
The hydrogels can be used for 3D culture. Faster relaxation led to increased spreading and proliferation of fibroblasts, and enhanced osteogenic differentiation of MSCs
3T3 fibroblasts; MSCs
74
Alginate interpenetrating network as an artificial ECM
Prepared by a combination of ionic and covalent cross-linking of click-functionalised alginate, interpenetrating with fibrillar collagen type I
Varying the mode and magnitude of crosslinking enables tuneable stiffness and viscoelasticity
MSC expression of immunomodulatory markers was differentially impacted by the viscoelasticity and stiffness of the matrix
MSCs
75
Boronate ester hydrogel
Prepared by reversible boronate esterification of boronic acid with vic-diols
Viscoelasticity increased as a function of the boronic acid and vicinal diol concentration, and also increased with decreasing cross-linker concentration, where the maximal loss tangent achieved was 0.55 at 0.1 rad/s
The cell area and nuclear area, focal adhesion tension, and subcellular location of YAP/TAZ were found to be lower for cells cultured on viscoelastic hydrogels compared to elastic hydrogels with a similar storage modulus
NIH-3T3 cells
78
Boronate-based hydrogels
Based on reversible boronate bonds
Relaxation time constants on the order of seconds or less
Fast relaxation matrix mechanics are found to promote cell-matrix interactions, leading to spreading and an increase in nuclear volume, and induce YAP/TAZ binding domain nuclear localization at longer times
MSCs
79
Collagen gels
Fabricated by adjusting pH
Strain-enhanced stress relaxation of collagen gels arises from force-dependent unbinding of weak bonds between collagen fibres
-
-
76
Hyaluronic acid hydrogels
Crosslinked via photo-responsive guest-host pairing of azobenzene to β-cyclodextrin
Relaxation time from 6 seconds to minutes
The hydrogels maintained a high level of viability after 3 days of culture
NIH 3T3 cells
77
Hyaluronic acid
Combined light-mediated covalent and supramolecular crosslinking was used to afford spatiotemporal control of the viscoelastic network
Significantly higher loss moduli compared to elastic group. Photopatterning enabled presentation of dynamic, heterogeneous viscoelastic properties
LX-2 cells respond to the viscoelastic hydrogel by displaying reductions in spread area, MRTF-A nuclear translation, and organisation of actin stress fibres
LX-2 stellate cells
80
Hyaluronic acid-collagen hydrogels
Interpenetrating network based on HA crosslinked with dynamic hydrazone bonds with collagen type I
The time for the initial stress of the material to be relaxed to half its value during a stress relaxation test (τ1/2) was modulated from ~233 seconds to > 18000 seconds
Faster relaxation promotes cell spreading, fibre remodelling, and focal adhesion formation in 3D culture
Human MSCs
81
Oxime cross-linked alginate hydrogels
Formed by mixing alkoxyamine-containing alginate with aldehyde-containing alginate
Stress-relaxation was tuneable by varying the composition or environmental factors
The gels showed very nice short-term cytocompatibility with the encapsulated cells. Growth and migration benefited from the stress relaxation capability
2PK3 cells
82
PEG hydrogels
Crosslinked by reversible hydrazone bonds
τ1/2 could be varied from 5–6000 seconds by changing the number of PEG or by changing the ratio of benzaldehyde to aliphatic aldehyde crosslinkers
Covalently-adaptable hydrogels allowed for the development of physiologically-relevant morphologies, whereas non-adaptable gels prevented cytoskeletal rearrangement and extension
C2C12 myoblasts
83
Thioester hydrogel
Photopolymerisation between PEG-SH and thioester-containing divinyl crosslinker
Through control of pH, gel stoichiometry, and crosslinker structure, viscoelastic properties were modulated across several orders of magnitude
MSCs encapsulated in the thioester hydrogels were able to elongate in 3D and display increased proliferation relative to those in static hydrogels
MSCs
84
Hyaluronic acid and PEG
A DN was formed based on the combination of supramolecular GH hyaluronic acid networks with covalent networks from the photocrosslinking of PEG-fibrinogen and PEG-diacrylate
Dependent on the polymer concentration the GH network
The increase of GH concentration led to the enhancement of the viscosity of the DN hydrogel and the enhancement of cell spreading and proliferation
Stress-relaxation behavior in gels with ionic and covalent crosslinks
1
2010
... Alginate, a natural material, has been used to fabricate viscoelastic matrices for cell culture. The viscoelasticity can be controlled by changing the ratio of ionic cross-linker to covalent cross-linker, whose stress relaxation occurrs as a result of crosslinker unbinding.86 Based on this, Chaudhuri’s group.73, 87 reported that alginate hydrogels with various stress relaxation properties were obtained by changing the molecular weight of alginate or covalent incorporation of short PEG chains, which modulated the density of ionic crosslinks or led to a looser network in gels. Most recently, the same group found that stress relaxation of alginate hydrogel was simply and precisely modulated by changing the molecular weight and concentration of PEG.74 Increasing PEG concentration and molecular weight led to faster stress relaxation, higher loss modulus and increased creep. In order to simulate the fibrillar architecture of natural ECM, an interpenetrating network of click-functionalised alginate and fibrillar collagen type I was fabricated as a viscoelastic artificial ECM.75 Timing sequential ionic and covalent crosslinking allowed collagen type I to self-assemble into fibrous structures within the network. The stiffness and viscoelasticity was tuned by changing the mode and magnitude of crosslinking, without changing the micro-scale architecture of the hydrogels. The above viscoelastic alginate hydrogels were mostly ionically cross-linked, mainly using physical approaches to regulate viscoelasticity. Besides the ionic interaction, other physically cross-linked hydrogels have also been explored to form viscoelastic hydrogels including hydrogen bonding and guest-host interaction. ...
Mechanical confinement regulates cartilage matrix formation by chondrocytes
2
2017
... Alginate, a natural material, has been used to fabricate viscoelastic matrices for cell culture. The viscoelasticity can be controlled by changing the ratio of ionic cross-linker to covalent cross-linker, whose stress relaxation occurrs as a result of crosslinker unbinding.86 Based on this, Chaudhuri’s group.73, 87 reported that alginate hydrogels with various stress relaxation properties were obtained by changing the molecular weight of alginate or covalent incorporation of short PEG chains, which modulated the density of ionic crosslinks or led to a looser network in gels. Most recently, the same group found that stress relaxation of alginate hydrogel was simply and precisely modulated by changing the molecular weight and concentration of PEG.74 Increasing PEG concentration and molecular weight led to faster stress relaxation, higher loss modulus and increased creep. In order to simulate the fibrillar architecture of natural ECM, an interpenetrating network of click-functionalised alginate and fibrillar collagen type I was fabricated as a viscoelastic artificial ECM.75 Timing sequential ionic and covalent crosslinking allowed collagen type I to self-assemble into fibrous structures within the network. The stiffness and viscoelasticity was tuned by changing the mode and magnitude of crosslinking, without changing the micro-scale architecture of the hydrogels. The above viscoelastic alginate hydrogels were mostly ionically cross-linked, mainly using physical approaches to regulate viscoelasticity. Besides the ionic interaction, other physically cross-linked hydrogels have also been explored to form viscoelastic hydrogels including hydrogen bonding and guest-host interaction. ...
... The viscoelastic properties of the ECM have been elucidated as a regulator of cell behaviours such as FA formation, cell spreading, proliferation, migration, and differentiation. Recently, there are some studies which showed the biological impacts of viscoelasticity in vivo. For example, Ghosh et al.92 prepared an ionically-crosslinked chitosan hydrogel with viscoelasticity to repair osteochondral defects in rabbits. They found that strain-reversible gels supported early cartilage-specific matrix formation and woven bone deposition. In a following study, the same group introduced stable covalent bonds into the gels to improve mechanical stability in vivo, and found that dual cross-linked chitosan hydrogels exhibited more collagen and mineral deposition compared to single ionically-crosslinked gels.93 The τ1/2 of a substrate has been proven to regulate many cell behaviours including stem cell differentiation in vitro. Mooney’s group addressed the hypothesis that substrate stress relaxation time could affect bone regeneration in vivo. They prepared alginate gels with various stress-relaxation times, which they then implanted into rat calvarial defects together with hMSCs.94 After 3 months of implantation, the rats that received fast-relaxing hydrogels (τ1/2 ≈ 50 seconds) showed much more growth of new bone compared with the rats that received slow-relaxing, stiffness-matched hydrogels. Strikingly, fast-relaxing hydrogel without encapsulated MSCs also promoted new bone formation, which was only slightly less than that induced by fast-relaxing hydrogels with MSCs (Figure 5). These results suggested that the viscoelasticity of biomaterials was an effective regulator of tissue regeneration in vivo, just like chemical and biological cues. Tissue engineering provides a promising solution for repair or regeneration of cartilage defects. To improve regenerative outcomes, matrices have been designed to incorporate viscoelastic features, making them much more similar to the mechanical features of native cartilage. To give one example, Lee et al.87 tested the impact of hydrogel stress relaxation rates on chondrocytes in 3D culture for cartilage tissue engineering. They found that faster relaxation promoted a significant increase in volume of interconnected cartilage matrix, while slower relaxation resulted in increased interleukin-1β secretion. In a recent study, Richardson et al.95 developed hydrazone crosslinked PEG hydrogels with tuneable viscoelastic properties for cartilage tissue engineering. They found that after 4 weeks of encapsulation, chondrocytes had more interconnected articular cartilage-specific matrix in stress-relaxation hydrogels. Collagen and sulphated glycosaminoglycans were deposited most in the adaptable hydrogel (22% benzyl-hydrazone hydrogel). Most recently, Li et al.96 used double network (DN) hydrogels based on gellan gum and polyethylene glycol acrylate (GG/PEGDA/DN) with stress-relaxing behaviour to study the effect on chondrogenic differentiation of BMSCs in a subcutaneous implant model. The results indicated that GG/PEGDA/DN provided a good physical environment for cell proliferation and spreading, and induced BMSCs to differentiate into the chondrogenic lineage, as demonstrated by a much greater level of collagen type II and transforming growth factor-β1. All these results show that stress relaxation of a substrate can mediate scaffold remodelling and tissue formation, which provides a new parameter to optimize tissue regeneration. ...
Insoluble elastin reduces collagen scaffold stiffness, improves viscoelastic properties, and induces a contractile phenotype in smooth muscle cells
1
2015
... The proliferation of various resident cells in a viscoelastic microenvironment was also found to be enhanced and regulated. Several studies demonstrated that hydrogel viscoelasticity has an obvious effect in promoting spreading and proliferation of fibroblasts in 2D or 3D culture.73, 88, 89 Similarly, when hMSCs were cultured on polyacrylamide-based viscoelastic hydrogels, the proliferation rate of hMSCs was significantly promoted by faster stress relaxation times.64 On an oxime cross-linked alginate hydrogel,82 2PK3 cells showed the same trend, with cells growing better in hydrogels with fast stress relaxation. All results indicate that stress relaxation time-dependent regulation may have beneficial effects on proliferation. ...
Hydrogel substrate stress-relaxation regulates the spreading and proliferation of mouse myoblasts
3
2017
... The proliferation of various resident cells in a viscoelastic microenvironment was also found to be enhanced and regulated. Several studies demonstrated that hydrogel viscoelasticity has an obvious effect in promoting spreading and proliferation of fibroblasts in 2D or 3D culture.73, 88, 89 Similarly, when hMSCs were cultured on polyacrylamide-based viscoelastic hydrogels, the proliferation rate of hMSCs was significantly promoted by faster stress relaxation times.64 On an oxime cross-linked alginate hydrogel,82 2PK3 cells showed the same trend, with cells growing better in hydrogels with fast stress relaxation. All results indicate that stress relaxation time-dependent regulation may have beneficial effects on proliferation. ...
... There is evidence that substrate stiffness guides cell migration, as does substrate viscoelasticity. Some studies have shown that mechanical cues from a viscoelastic matrix provide the orientation for migration. For instance, epithelial monolayer cells showed a special coalescence on a viscoelastic substrate, which was not observed on elastic materials.90 Sarcoma cells showed a faster migration speed in a collagen-based hypoxic hydrogel with faster stress relaxation than that in a gel with lower stress relaxation.89 Although it has been reported that cell migration can be regulated by the viscoelasticity of 2D or 3D substrates, the mechanism of this process needs to be further elucidated. ...
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
Epithelial monolayers coalesce on a viscoelastic substrate through redistribution of vinculin
1
2017
... There is evidence that substrate stiffness guides cell migration, as does substrate viscoelasticity. Some studies have shown that mechanical cues from a viscoelastic matrix provide the orientation for migration. For instance, epithelial monolayer cells showed a special coalescence on a viscoelastic substrate, which was not observed on elastic materials.90 Sarcoma cells showed a faster migration speed in a collagen-based hypoxic hydrogel with faster stress relaxation than that in a gel with lower stress relaxation.89 Although it has been reported that cell migration can be regulated by the viscoelasticity of 2D or 3D substrates, the mechanism of this process needs to be further elucidated. ...
Time to relax: mechanical stress release guides stem cell responses
1
2016
... Stem cells have similar reactions to viscoelasticity in terms of cell spreading, proliferation, and migration. However, stem cells are unique in development and differentiation. Viscoelasticity has gained researchers’ attention recently for its role in directing stem-cell fate,14, 91 as well as elasticity emphasized in cell differentiation. ...
Citrate cross-linked gels with strain reversibility and viscoelastic behavior accelerate healing of osteochondral defects in a rabbit model
1
2014
... The viscoelastic properties of the ECM have been elucidated as a regulator of cell behaviours such as FA formation, cell spreading, proliferation, migration, and differentiation. Recently, there are some studies which showed the biological impacts of viscoelasticity in vivo. For example, Ghosh et al.92 prepared an ionically-crosslinked chitosan hydrogel with viscoelasticity to repair osteochondral defects in rabbits. They found that strain-reversible gels supported early cartilage-specific matrix formation and woven bone deposition. In a following study, the same group introduced stable covalent bonds into the gels to improve mechanical stability in vivo, and found that dual cross-linked chitosan hydrogels exhibited more collagen and mineral deposition compared to single ionically-crosslinked gels.93 The τ1/2 of a substrate has been proven to regulate many cell behaviours including stem cell differentiation in vitro. Mooney’s group addressed the hypothesis that substrate stress relaxation time could affect bone regeneration in vivo. They prepared alginate gels with various stress-relaxation times, which they then implanted into rat calvarial defects together with hMSCs.94 After 3 months of implantation, the rats that received fast-relaxing hydrogels (τ1/2 ≈ 50 seconds) showed much more growth of new bone compared with the rats that received slow-relaxing, stiffness-matched hydrogels. Strikingly, fast-relaxing hydrogel without encapsulated MSCs also promoted new bone formation, which was only slightly less than that induced by fast-relaxing hydrogels with MSCs (Figure 5). These results suggested that the viscoelasticity of biomaterials was an effective regulator of tissue regeneration in vivo, just like chemical and biological cues. Tissue engineering provides a promising solution for repair or regeneration of cartilage defects. To improve regenerative outcomes, matrices have been designed to incorporate viscoelastic features, making them much more similar to the mechanical features of native cartilage. To give one example, Lee et al.87 tested the impact of hydrogel stress relaxation rates on chondrocytes in 3D culture for cartilage tissue engineering. They found that faster relaxation promoted a significant increase in volume of interconnected cartilage matrix, while slower relaxation resulted in increased interleukin-1β secretion. In a recent study, Richardson et al.95 developed hydrazone crosslinked PEG hydrogels with tuneable viscoelastic properties for cartilage tissue engineering. They found that after 4 weeks of encapsulation, chondrocytes had more interconnected articular cartilage-specific matrix in stress-relaxation hydrogels. Collagen and sulphated glycosaminoglycans were deposited most in the adaptable hydrogel (22% benzyl-hydrazone hydrogel). Most recently, Li et al.96 used double network (DN) hydrogels based on gellan gum and polyethylene glycol acrylate (GG/PEGDA/DN) with stress-relaxing behaviour to study the effect on chondrogenic differentiation of BMSCs in a subcutaneous implant model. The results indicated that GG/PEGDA/DN provided a good physical environment for cell proliferation and spreading, and induced BMSCs to differentiate into the chondrogenic lineage, as demonstrated by a much greater level of collagen type II and transforming growth factor-β1. All these results show that stress relaxation of a substrate can mediate scaffold remodelling and tissue formation, which provides a new parameter to optimize tissue regeneration. ...
Covalent cross-links in polyampholytic chitosan fibers enhances bone regeneration in a rabbit model
1
2015
... The viscoelastic properties of the ECM have been elucidated as a regulator of cell behaviours such as FA formation, cell spreading, proliferation, migration, and differentiation. Recently, there are some studies which showed the biological impacts of viscoelasticity in vivo. For example, Ghosh et al.92 prepared an ionically-crosslinked chitosan hydrogel with viscoelasticity to repair osteochondral defects in rabbits. They found that strain-reversible gels supported early cartilage-specific matrix formation and woven bone deposition. In a following study, the same group introduced stable covalent bonds into the gels to improve mechanical stability in vivo, and found that dual cross-linked chitosan hydrogels exhibited more collagen and mineral deposition compared to single ionically-crosslinked gels.93 The τ1/2 of a substrate has been proven to regulate many cell behaviours including stem cell differentiation in vitro. Mooney’s group addressed the hypothesis that substrate stress relaxation time could affect bone regeneration in vivo. They prepared alginate gels with various stress-relaxation times, which they then implanted into rat calvarial defects together with hMSCs.94 After 3 months of implantation, the rats that received fast-relaxing hydrogels (τ1/2 ≈ 50 seconds) showed much more growth of new bone compared with the rats that received slow-relaxing, stiffness-matched hydrogels. Strikingly, fast-relaxing hydrogel without encapsulated MSCs also promoted new bone formation, which was only slightly less than that induced by fast-relaxing hydrogels with MSCs (Figure 5). These results suggested that the viscoelasticity of biomaterials was an effective regulator of tissue regeneration in vivo, just like chemical and biological cues. Tissue engineering provides a promising solution for repair or regeneration of cartilage defects. To improve regenerative outcomes, matrices have been designed to incorporate viscoelastic features, making them much more similar to the mechanical features of native cartilage. To give one example, Lee et al.87 tested the impact of hydrogel stress relaxation rates on chondrocytes in 3D culture for cartilage tissue engineering. They found that faster relaxation promoted a significant increase in volume of interconnected cartilage matrix, while slower relaxation resulted in increased interleukin-1β secretion. In a recent study, Richardson et al.95 developed hydrazone crosslinked PEG hydrogels with tuneable viscoelastic properties for cartilage tissue engineering. They found that after 4 weeks of encapsulation, chondrocytes had more interconnected articular cartilage-specific matrix in stress-relaxation hydrogels. Collagen and sulphated glycosaminoglycans were deposited most in the adaptable hydrogel (22% benzyl-hydrazone hydrogel). Most recently, Li et al.96 used double network (DN) hydrogels based on gellan gum and polyethylene glycol acrylate (GG/PEGDA/DN) with stress-relaxing behaviour to study the effect on chondrogenic differentiation of BMSCs in a subcutaneous implant model. The results indicated that GG/PEGDA/DN provided a good physical environment for cell proliferation and spreading, and induced BMSCs to differentiate into the chondrogenic lineage, as demonstrated by a much greater level of collagen type II and transforming growth factor-β1. All these results show that stress relaxation of a substrate can mediate scaffold remodelling and tissue formation, which provides a new parameter to optimize tissue regeneration. ...
Substrate stress-relaxation regulates scaffold remodeling and bone formation in vivo
2
2017
... The viscoelastic properties of the ECM have been elucidated as a regulator of cell behaviours such as FA formation, cell spreading, proliferation, migration, and differentiation. Recently, there are some studies which showed the biological impacts of viscoelasticity in vivo. For example, Ghosh et al.92 prepared an ionically-crosslinked chitosan hydrogel with viscoelasticity to repair osteochondral defects in rabbits. They found that strain-reversible gels supported early cartilage-specific matrix formation and woven bone deposition. In a following study, the same group introduced stable covalent bonds into the gels to improve mechanical stability in vivo, and found that dual cross-linked chitosan hydrogels exhibited more collagen and mineral deposition compared to single ionically-crosslinked gels.93 The τ1/2 of a substrate has been proven to regulate many cell behaviours including stem cell differentiation in vitro. Mooney’s group addressed the hypothesis that substrate stress relaxation time could affect bone regeneration in vivo. They prepared alginate gels with various stress-relaxation times, which they then implanted into rat calvarial defects together with hMSCs.94 After 3 months of implantation, the rats that received fast-relaxing hydrogels (τ1/2 ≈ 50 seconds) showed much more growth of new bone compared with the rats that received slow-relaxing, stiffness-matched hydrogels. Strikingly, fast-relaxing hydrogel without encapsulated MSCs also promoted new bone formation, which was only slightly less than that induced by fast-relaxing hydrogels with MSCs (Figure 5). These results suggested that the viscoelasticity of biomaterials was an effective regulator of tissue regeneration in vivo, just like chemical and biological cues. Tissue engineering provides a promising solution for repair or regeneration of cartilage defects. To improve regenerative outcomes, matrices have been designed to incorporate viscoelastic features, making them much more similar to the mechanical features of native cartilage. To give one example, Lee et al.87 tested the impact of hydrogel stress relaxation rates on chondrocytes in 3D culture for cartilage tissue engineering. They found that faster relaxation promoted a significant increase in volume of interconnected cartilage matrix, while slower relaxation resulted in increased interleukin-1β secretion. In a recent study, Richardson et al.95 developed hydrazone crosslinked PEG hydrogels with tuneable viscoelastic properties for cartilage tissue engineering. They found that after 4 weeks of encapsulation, chondrocytes had more interconnected articular cartilage-specific matrix in stress-relaxation hydrogels. Collagen and sulphated glycosaminoglycans were deposited most in the adaptable hydrogel (22% benzyl-hydrazone hydrogel). Most recently, Li et al.96 used double network (DN) hydrogels based on gellan gum and polyethylene glycol acrylate (GG/PEGDA/DN) with stress-relaxing behaviour to study the effect on chondrogenic differentiation of BMSCs in a subcutaneous implant model. The results indicated that GG/PEGDA/DN provided a good physical environment for cell proliferation and spreading, and induced BMSCs to differentiate into the chondrogenic lineage, as demonstrated by a much greater level of collagen type II and transforming growth factor-β1. All these results show that stress relaxation of a substrate can mediate scaffold remodelling and tissue formation, which provides a new parameter to optimize tissue regeneration. ...
... 94 Copyright Wiley-VCH Verlag GmbH & Co. KGaA. Comparison of Cellular Modulation Mediated by Elasticity and Viscoelasticity
In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
... The viscoelastic properties of the ECM have been elucidated as a regulator of cell behaviours such as FA formation, cell spreading, proliferation, migration, and differentiation. Recently, there are some studies which showed the biological impacts of viscoelasticity in vivo. For example, Ghosh et al.92 prepared an ionically-crosslinked chitosan hydrogel with viscoelasticity to repair osteochondral defects in rabbits. They found that strain-reversible gels supported early cartilage-specific matrix formation and woven bone deposition. In a following study, the same group introduced stable covalent bonds into the gels to improve mechanical stability in vivo, and found that dual cross-linked chitosan hydrogels exhibited more collagen and mineral deposition compared to single ionically-crosslinked gels.93 The τ1/2 of a substrate has been proven to regulate many cell behaviours including stem cell differentiation in vitro. Mooney’s group addressed the hypothesis that substrate stress relaxation time could affect bone regeneration in vivo. They prepared alginate gels with various stress-relaxation times, which they then implanted into rat calvarial defects together with hMSCs.94 After 3 months of implantation, the rats that received fast-relaxing hydrogels (τ1/2 ≈ 50 seconds) showed much more growth of new bone compared with the rats that received slow-relaxing, stiffness-matched hydrogels. Strikingly, fast-relaxing hydrogel without encapsulated MSCs also promoted new bone formation, which was only slightly less than that induced by fast-relaxing hydrogels with MSCs (Figure 5). These results suggested that the viscoelasticity of biomaterials was an effective regulator of tissue regeneration in vivo, just like chemical and biological cues. Tissue engineering provides a promising solution for repair or regeneration of cartilage defects. To improve regenerative outcomes, matrices have been designed to incorporate viscoelastic features, making them much more similar to the mechanical features of native cartilage. To give one example, Lee et al.87 tested the impact of hydrogel stress relaxation rates on chondrocytes in 3D culture for cartilage tissue engineering. They found that faster relaxation promoted a significant increase in volume of interconnected cartilage matrix, while slower relaxation resulted in increased interleukin-1β secretion. In a recent study, Richardson et al.95 developed hydrazone crosslinked PEG hydrogels with tuneable viscoelastic properties for cartilage tissue engineering. They found that after 4 weeks of encapsulation, chondrocytes had more interconnected articular cartilage-specific matrix in stress-relaxation hydrogels. Collagen and sulphated glycosaminoglycans were deposited most in the adaptable hydrogel (22% benzyl-hydrazone hydrogel). Most recently, Li et al.96 used double network (DN) hydrogels based on gellan gum and polyethylene glycol acrylate (GG/PEGDA/DN) with stress-relaxing behaviour to study the effect on chondrogenic differentiation of BMSCs in a subcutaneous implant model. The results indicated that GG/PEGDA/DN provided a good physical environment for cell proliferation and spreading, and induced BMSCs to differentiate into the chondrogenic lineage, as demonstrated by a much greater level of collagen type II and transforming growth factor-β1. All these results show that stress relaxation of a substrate can mediate scaffold remodelling and tissue formation, which provides a new parameter to optimize tissue regeneration. ...
Stress-relaxing double-network hydrogel for chondrogenic differentiation of stem cells
1
2020
... The viscoelastic properties of the ECM have been elucidated as a regulator of cell behaviours such as FA formation, cell spreading, proliferation, migration, and differentiation. Recently, there are some studies which showed the biological impacts of viscoelasticity in vivo. For example, Ghosh et al.92 prepared an ionically-crosslinked chitosan hydrogel with viscoelasticity to repair osteochondral defects in rabbits. They found that strain-reversible gels supported early cartilage-specific matrix formation and woven bone deposition. In a following study, the same group introduced stable covalent bonds into the gels to improve mechanical stability in vivo, and found that dual cross-linked chitosan hydrogels exhibited more collagen and mineral deposition compared to single ionically-crosslinked gels.93 The τ1/2 of a substrate has been proven to regulate many cell behaviours including stem cell differentiation in vitro. Mooney’s group addressed the hypothesis that substrate stress relaxation time could affect bone regeneration in vivo. They prepared alginate gels with various stress-relaxation times, which they then implanted into rat calvarial defects together with hMSCs.94 After 3 months of implantation, the rats that received fast-relaxing hydrogels (τ1/2 ≈ 50 seconds) showed much more growth of new bone compared with the rats that received slow-relaxing, stiffness-matched hydrogels. Strikingly, fast-relaxing hydrogel without encapsulated MSCs also promoted new bone formation, which was only slightly less than that induced by fast-relaxing hydrogels with MSCs (Figure 5). These results suggested that the viscoelasticity of biomaterials was an effective regulator of tissue regeneration in vivo, just like chemical and biological cues. Tissue engineering provides a promising solution for repair or regeneration of cartilage defects. To improve regenerative outcomes, matrices have been designed to incorporate viscoelastic features, making them much more similar to the mechanical features of native cartilage. To give one example, Lee et al.87 tested the impact of hydrogel stress relaxation rates on chondrocytes in 3D culture for cartilage tissue engineering. They found that faster relaxation promoted a significant increase in volume of interconnected cartilage matrix, while slower relaxation resulted in increased interleukin-1β secretion. In a recent study, Richardson et al.95 developed hydrazone crosslinked PEG hydrogels with tuneable viscoelastic properties for cartilage tissue engineering. They found that after 4 weeks of encapsulation, chondrocytes had more interconnected articular cartilage-specific matrix in stress-relaxation hydrogels. Collagen and sulphated glycosaminoglycans were deposited most in the adaptable hydrogel (22% benzyl-hydrazone hydrogel). Most recently, Li et al.96 used double network (DN) hydrogels based on gellan gum and polyethylene glycol acrylate (GG/PEGDA/DN) with stress-relaxing behaviour to study the effect on chondrogenic differentiation of BMSCs in a subcutaneous implant model. The results indicated that GG/PEGDA/DN provided a good physical environment for cell proliferation and spreading, and induced BMSCs to differentiate into the chondrogenic lineage, as demonstrated by a much greater level of collagen type II and transforming growth factor-β1. All these results show that stress relaxation of a substrate can mediate scaffold remodelling and tissue formation, which provides a new parameter to optimize tissue regeneration. ...
Opposite responses of normal hepatocytes and hepatocellular carcinoma cells to substrate viscoelasticity
1
2020
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
Nanoscale viscoelastic properties and adhesion of polydimethylsiloxane for tissue engineering
1
2014
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
Interplay of matrix stiffness and protein tethering in stem cell differentiation
1
2014
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
Divergent immune responses to synthetic and biological scaffolds
1
2019
... In order to clearly understand the effect of elastic and viscoelastic materials on cells, it may be necessary to make a direct comparison between the two types of materials. Chaudhuri et al.72 compared the effect of viscoelasticity and linear elasticity on U2OS cell spreading at various initial elastic moduli (1.4, 3.4 and 9 kPa) with ionically-crosslinked and covalently-crosslinked alginate hydrogels. At lower moduli (1.4 kPa), greater cell spreading was observed on viscoelastic substrates compared to the purely elastic substrates, and the effect was enhanced with increasing ligand density. Further investigation revealed that the effect of viscoelastic substrates at low initial elastic moduli on cell spreading was mediated by integrin-β1, actin aggregation, actomyosin contraction and increased nuclear localization with yes-associated protein (YAP). In a similar way, Bauer et al.89 used alginate hydrogels to explore the effect of viscoelasticity on the spreading and proliferation of mouse myoblasts at various initial elastic moduli relative to linear elasticity. At lower initial elastic moduli (2.8 and 12.2 kPa), cell spreading on the viscoelastic substrates was greater than that on purely elastic substrates, while the opposite result was observed at higher initial elastic moduli (49.5 kPa). Compared with elastic substrates, myoblasts on viscoelastic substrates were more elongated, and this effect was enhanced as the modulus increased. Moreover, cell proliferation on viscoelastic substrates was greater than on purely elastic substrates. In addition to the initial elastic modulus, the influence of viscoelasticity and elasticity on cell spreading may also be influenced by the different cell types. Mandal et al.97 found that the spreading area and actin bundle assembly of primary human hepatocytes and fibroblasts were reduced on viscoelastic substrates compared to purely elastic substrates with the same stiffness. However, Huh7 cells spread faster on viscoelastic substrates than on substrates with linear elasticity. Polyacrylamide hydrogels have been widely used to study the effect of substrate elasticity on cells due to their easy availability and formulation stability. More recently, Charrier et al.71 prepared purely elastic and viscoelastic hydrogels by using a mixture of polyacrylamide and linear polyacrylamide, and studied their effects on cell behaviour. They found that the cell area on the viscoelastic hydrogel was smaller than that on the purely elastic hydrogel at the same storage modulus. Hydrogels with the same elasticity and storage moduli but coated with either collagen or fibronectin showed different cell spreading, suggesting that ligands also varied the effect of the viscoelasticity on cells. Moreover, viscoelastic hydrogels resulted in a reduced differentiation ratio of rat hepatic stellate cells to myofibroblasts compared with purely elastic hydrogels. Interestingly, recovery experiments confirmed that viscoelasticity also promoted the dedifferentiation of established myofibroblasts. PDMS has often been regarded as another common elastic material for studying cell behaviour.18 However, it was found that many formulations of PDMS are viscoelastic at the nano (cell) scale, which may be due to the use of different mechanical characterisation methods.98 This apparent mismatch might lead to different cellular responses to the mechanical properties of the substrate to those expected.99, 100 Therefore, when using PDMS to study the effects of mechanical cues on cell behaviour, it is necessary to take into account the differences in the mechanical properties of materials at the macroscopic and nano scale. Recently, the work of Sadtler et al.101 may provide new insights into the regulation of viscoelasticity and elasticity on cell fate. Compared with viscoelastic natural ECM, purely elastic synthetic matrix recruits more neutrophils leading to inflammation rather than promoting regenerative immune cell responses. To date, there have been few studies on the direct comparison between purely elastic and viscoelastic materials. More exploration in this field is necessary in the future so as to more clearly explain the regulation of these two mechanical cues on cell behaviours. ...
Substrate stiffness- and topography-dependent differentiation of annulus fibrosus-derived stem cells is regulated by Yes-associated protein
4
2019
... A growing number of studies have shown that cell fate can be directed by the stiffness or viscoelasticity of the ECM. However, the ECM of various natural tissues such as bone, nerves, skin, kidney, or AF, has distinct topography at nano- or micro-scales.10, 102-105 Topographic factors including size, shape, and geometric arrangement, also considered to be an important biophysical cue, have substantial influence on cell behaviours. ...
... Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
... In addition, the stiffness of fibrous scaffolds also plays an important role in disease development processes or tissue repair processes. For example, our group recently reported that fibre stiffness of polyurethanes and fibre diameters showed a synergistic effect on the differentiation of AFSCs. AFSCs had a strong tendency to differentiate into various regional AF-like cells when cultured on a scaffold with physical cues similar to native AF tissue.102 Davidson et al.118 seeded hepatic stellate cells onto soft or stiff fibrous hyaluronic acid hydrogels, and found that cell spreading, α-smooth muscle actin expression, and formation of multicellular clusters decreased on stiff fibres compared with those on soft fibres, indicating the important influence of fibre stiffness on cell-material interactions and implying its relevance to disease and tissue repair processes. ...
... Commonly-used biomaterials with various elasticities and their effects on cells.
Material
Fabrication method
Elasticity
Effects on cell behaviours
Cell source
Reference
Alginate
Alginate microspheres prepared by microfluidic technology with different elasticities and microarchitectures,controlled by calcium ion concentrations.
18, 32 kPa
Elasticity and porosity regulated the fate of encapsulated MSCs through modulation of the nuclear factor-κB pathway
MSCs
34
Chitosan-hyaluronic acid
Porous chitosan-hyaluronic acid scaffolds of varied stiffness were fabricated using a phase separation method
1.41–27.7 kPa
Increased matrix stiffness resulted in increased drug resistance of glioblastoma multiforme cells, and elevated expression of drug resistance-,hypoxia-, and invasion-related genes
Glioblastoma multiforme cells
21
Dynamic protein hydrogels
A Ru2+-mediated photochemical strategy was used to crosslink an aqueous solution of FGR(G-MEP-R)2 into a chemically-crosslinked protein hydrogel
6–20 kPa
Human lung fibroblasts dynamically responded to changes of hydrogel mechanics in a reversible fashion, regulated by redox state
Human lung fibroblasts
39
Fibrin-alginate
Mechanical properties were tuneable via calcium chloride crosslinking
0.6–3.8 kPa
Spreading of MSCs and endothelial cells was a function of alginate crosslinking density
MSCs, endothelial cells
22
Hyaluronic acid
Methacrylated hyaluronic acid was synthesized to allow for crosslinking via Michael addition using the crosslinker dithiothreitol
0.2–4.5 kPa
Human breast cancer cell (MDA-MB-231Br) adhesion, spreading, proliferation and migration were tightly regulated by the hydrogel stiffness
MDA-MB-231Br
23
Polyacrylamide
Stiffness of polyacrylamide gels was adjusted using different monomer-to-crosslinker formulations
2–32 kPa
Cytoskeleton assembly and cell morphology were efficiently regulated by substrate stiffness
HeLa cells
24
Poly (dimethylsiloxane)
Poly(dimethylsiloxane) was used as the base material in which iron particles were embedded to create a magnetorheological elastomer, whose elasticity was controlled by the spacer distances between the magnet and the samples
10–55 kPa
The softer substrates yielded more organised sarcomeres,and sarcomere formation was positively correlated with the degree of myocyte enrichment when using human-derived induced pluripotent stem cell cardiomyocytes
Controlling the crosslinking of tri-block copolymer and polycaprolactone triol yielded polyurethanes of varying elasticity
45.0–244.8 kPa
Scaffolds with different stiffnesses stimulated the proliferation of different types of cells
3T3 fibroblasts, MG63 cells
26
Silk fibroin
Developed by introducing inert silk fibroin nanofibres within an enzyme crosslinked system of silk fibroin
9–60 kPa
MSCs differentiated into endothelial, myoblast and osteoblast cells on the different elastic substrates
MSCs
35
Silk fibroin-collagen
The concentrations of both proteins was changed gradually while maintaining the ratio at 1:7, which resulted in a gradual change in stiffness at a fixed composition
0.1–20 kPa
High rigidity allowed human MSCs to preserve all-directional spreading with polygonal shape. Soft substrates might not maintain the polygonal shape
Human MSCs
27
Poly(ether carbonate urethane)urea
Young’s modulus of scaffolds was tuned by adjusting the molecular weight of polydiol (soft segment) as well as the feed ratios of hard molecular segment to soft molecular segment
2.5–13.4 MPa
Annulus fibrosus-derived stem cells showed strong tendencies to differentiate into various types of annulus fibrosus-like cells depending on the substrate elasticity
Annulus fibrosus-derived stem cells
36, 102
PEG
Stiffness was adjusted by adding various PEG monomers and the photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate
1.5–12.6 kPa
The functional and molecular outputs of adult mouse ventricular myocytes were dependent on the PEG hydrogel stiffness
Fibre stiffness was controlled by altering the flow rates of the poly(L-lactic acid)-core and poly(L-lactide-co-caprolactone)-shell solutions.
14.7–2141.7 MPa
Higher stiffness of the aligned fibrous substrates was found to significantly encourage the proliferation and migration of human umbilical artery smooth muscle cells
Human umbilical arterial smooth muscle cells
29
GelMA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness by varying the concentration
3–180 kPa
PC12 cell viability, adhesion, spreading and average neurite length were influenced by stiffness
PC12 cells
30
GelMA/PEGDA hydrogels
Prepared by photocrosslinking methacrylate gelatine and adjusting the stiffness with the crosslinker PEGDA
4, 40 kPa
Increased matrix stiffness promoted osteogenic differentiation of MSCs
MSCs
31
GelMA/Collagen hydrogels
Prepared by mixing collagen and GelMA to form an interpenetrating network
2–12 kPa
With the increase of matrix stiffness, the invasion and sprouting of the two cells decreased regardless of fibre content
MDA-MB-231Br and endothelial cells
32
Alginate/GelMA hydrogels
Prepared by mixing alginate and GelMA
6–13 kPa
The expression level of MSC osteogenesis markers was enhanced with the increase in the matrix elastic modulus
Unusual inflammatory and fibrogenic pulmonary responses to single-walled carbon nanotubes in mice
1
2005
... Among these factors, size appears to play a critical role in modulating cell behaviours. For example, Nguyen et al.105 provided a comprehensive survey on the significance of its effect on cell adhesion, cell morphology and alignment, and neurite guidance. At both the nano- and micro-scale, cell behaviours can be significantly influenced by the size of topographical features. At the same time, the response of cells is diverse and strongly dependent on the type of cells. Cell behaviours often occur at both the nano- and micro-scales in biological interfaces and natural tissues, so an ideal artificial ECM should be biomimetic to match both size-scales and thus achieve a superior effect. For instance, ECs exhibited enhanced elongation when cultured on oriented nanofibrous scaffolds with different micro-grooved topographies.103 Recently, a 3D biomimetic nano-micro fibrous woven fabric/hydrogel composite scaffold was developed for heart valve engineering.106 The developers found that the native-like anisotropic features offered appropriate microenvironmental cues for cell proliferation and ECM deposition, representing a promising strategy to treat heart valve injury. ...
Emphysema and mechanical stress-induced lung remodeling
0
2013
From nano to micro: topographical scale and its impact on cell adhesion, morphology and contact guidance
4
2016
... A growing number of studies have shown that cell fate can be directed by the stiffness or viscoelasticity of the ECM. However, the ECM of various natural tissues such as bone, nerves, skin, kidney, or AF, has distinct topography at nano- or micro-scales.10, 102-105 Topographic factors including size, shape, and geometric arrangement, also considered to be an important biophysical cue, have substantial influence on cell behaviours. ...
... Among these factors, size appears to play a critical role in modulating cell behaviours. For example, Nguyen et al.105 provided a comprehensive survey on the significance of its effect on cell adhesion, cell morphology and alignment, and neurite guidance. At both the nano- and micro-scale, cell behaviours can be significantly influenced by the size of topographical features. At the same time, the response of cells is diverse and strongly dependent on the type of cells. Cell behaviours often occur at both the nano- and micro-scales in biological interfaces and natural tissues, so an ideal artificial ECM should be biomimetic to match both size-scales and thus achieve a superior effect. For instance, ECs exhibited enhanced elongation when cultured on oriented nanofibrous scaffolds with different micro-grooved topographies.103 Recently, a 3D biomimetic nano-micro fibrous woven fabric/hydrogel composite scaffold was developed for heart valve engineering.106 The developers found that the native-like anisotropic features offered appropriate microenvironmental cues for cell proliferation and ECM deposition, representing a promising strategy to treat heart valve injury. ...
... Alignment, one of the geometric arrangements, also exerts a great effect on cell behaviours. For cell adhesion, it was found that FAK phosphorylation was significantly increased in MSCs on aligned nanofibres compared with random nanofibres.105 For cell spreading, it was found that the cytoskeleton of human umbilical vein ECs on aligned fibres was stretched along the longitudinal direction of the fibre, while it extended in all directions on non-aligned fibres.107 Interestingly, cell alignment showed a more significant effect on micro-aligned scaffolds than on nano-aligned scaffolds.106 For cell migration, compared with random fibres, human adipose-derived stem cells cultured on aligned nanofibres exhibited a faster migration rate and higher synthetic ability of critical ECM molecules, such as collagen type I, tropoelastin, and matrix metalloproteinase-1.108 The effect of the alignment of electrospun polycaprolactone-gelatine ultrafine fibres on cell migration was also studied, and showed that the cell migration rate was faster in the direction of aligned fibres compared with non-aligned fibres.109 In addition, alignment affected cell proliferation and differentiation and was cell-type dependent. For example, aligned scaffold enhanced cell proliferation of keratocytes, while randomly-oriented scaffold enhanced the proliferation of corneal epithelial cells.110 The osteogenic differentiation of MSCs could also be modulated by fibre alignment. Alkaline phosphatase and calcium nodules were evenly distributed along the direction of aligned fibres, while gene expression of osteocalcin and osteopontin was significantly enhanced, indicating that aligned fibres were superior to non-aligned fibres in inducing osteogenic differentiation.107 ...
... Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
Living nano-micro fibrous woven fabric/hydrogel composite scaffolds for heart valve engineering
2
2017
... Among these factors, size appears to play a critical role in modulating cell behaviours. For example, Nguyen et al.105 provided a comprehensive survey on the significance of its effect on cell adhesion, cell morphology and alignment, and neurite guidance. At both the nano- and micro-scale, cell behaviours can be significantly influenced by the size of topographical features. At the same time, the response of cells is diverse and strongly dependent on the type of cells. Cell behaviours often occur at both the nano- and micro-scales in biological interfaces and natural tissues, so an ideal artificial ECM should be biomimetic to match both size-scales and thus achieve a superior effect. For instance, ECs exhibited enhanced elongation when cultured on oriented nanofibrous scaffolds with different micro-grooved topographies.103 Recently, a 3D biomimetic nano-micro fibrous woven fabric/hydrogel composite scaffold was developed for heart valve engineering.106 The developers found that the native-like anisotropic features offered appropriate microenvironmental cues for cell proliferation and ECM deposition, representing a promising strategy to treat heart valve injury. ...
... Alignment, one of the geometric arrangements, also exerts a great effect on cell behaviours. For cell adhesion, it was found that FAK phosphorylation was significantly increased in MSCs on aligned nanofibres compared with random nanofibres.105 For cell spreading, it was found that the cytoskeleton of human umbilical vein ECs on aligned fibres was stretched along the longitudinal direction of the fibre, while it extended in all directions on non-aligned fibres.107 Interestingly, cell alignment showed a more significant effect on micro-aligned scaffolds than on nano-aligned scaffolds.106 For cell migration, compared with random fibres, human adipose-derived stem cells cultured on aligned nanofibres exhibited a faster migration rate and higher synthetic ability of critical ECM molecules, such as collagen type I, tropoelastin, and matrix metalloproteinase-1.108 The effect of the alignment of electrospun polycaprolactone-gelatine ultrafine fibres on cell migration was also studied, and showed that the cell migration rate was faster in the direction of aligned fibres compared with non-aligned fibres.109 In addition, alignment affected cell proliferation and differentiation and was cell-type dependent. For example, aligned scaffold enhanced cell proliferation of keratocytes, while randomly-oriented scaffold enhanced the proliferation of corneal epithelial cells.110 The osteogenic differentiation of MSCs could also be modulated by fibre alignment. Alkaline phosphatase and calcium nodules were evenly distributed along the direction of aligned fibres, while gene expression of osteocalcin and osteopontin was significantly enhanced, indicating that aligned fibres were superior to non-aligned fibres in inducing osteogenic differentiation.107 ...
Effects of fiber alignment on stem cells-fibrous scaffold interactions
2
2015
... Alignment, one of the geometric arrangements, also exerts a great effect on cell behaviours. For cell adhesion, it was found that FAK phosphorylation was significantly increased in MSCs on aligned nanofibres compared with random nanofibres.105 For cell spreading, it was found that the cytoskeleton of human umbilical vein ECs on aligned fibres was stretched along the longitudinal direction of the fibre, while it extended in all directions on non-aligned fibres.107 Interestingly, cell alignment showed a more significant effect on micro-aligned scaffolds than on nano-aligned scaffolds.106 For cell migration, compared with random fibres, human adipose-derived stem cells cultured on aligned nanofibres exhibited a faster migration rate and higher synthetic ability of critical ECM molecules, such as collagen type I, tropoelastin, and matrix metalloproteinase-1.108 The effect of the alignment of electrospun polycaprolactone-gelatine ultrafine fibres on cell migration was also studied, and showed that the cell migration rate was faster in the direction of aligned fibres compared with non-aligned fibres.109 In addition, alignment affected cell proliferation and differentiation and was cell-type dependent. For example, aligned scaffold enhanced cell proliferation of keratocytes, while randomly-oriented scaffold enhanced the proliferation of corneal epithelial cells.110 The osteogenic differentiation of MSCs could also be modulated by fibre alignment. Alkaline phosphatase and calcium nodules were evenly distributed along the direction of aligned fibres, while gene expression of osteocalcin and osteopontin was significantly enhanced, indicating that aligned fibres were superior to non-aligned fibres in inducing osteogenic differentiation.107 ...
... 107 ...
Spatial arrangement of polycaprolactone/collagen nanofiber scaffolds regulates the wound healing related behaviors of human adipose stromal cells
1
2012
... Alignment, one of the geometric arrangements, also exerts a great effect on cell behaviours. For cell adhesion, it was found that FAK phosphorylation was significantly increased in MSCs on aligned nanofibres compared with random nanofibres.105 For cell spreading, it was found that the cytoskeleton of human umbilical vein ECs on aligned fibres was stretched along the longitudinal direction of the fibre, while it extended in all directions on non-aligned fibres.107 Interestingly, cell alignment showed a more significant effect on micro-aligned scaffolds than on nano-aligned scaffolds.106 For cell migration, compared with random fibres, human adipose-derived stem cells cultured on aligned nanofibres exhibited a faster migration rate and higher synthetic ability of critical ECM molecules, such as collagen type I, tropoelastin, and matrix metalloproteinase-1.108 The effect of the alignment of electrospun polycaprolactone-gelatine ultrafine fibres on cell migration was also studied, and showed that the cell migration rate was faster in the direction of aligned fibres compared with non-aligned fibres.109 In addition, alignment affected cell proliferation and differentiation and was cell-type dependent. For example, aligned scaffold enhanced cell proliferation of keratocytes, while randomly-oriented scaffold enhanced the proliferation of corneal epithelial cells.110 The osteogenic differentiation of MSCs could also be modulated by fibre alignment. Alkaline phosphatase and calcium nodules were evenly distributed along the direction of aligned fibres, while gene expression of osteocalcin and osteopontin was significantly enhanced, indicating that aligned fibres were superior to non-aligned fibres in inducing osteogenic differentiation.107 ...
Combined nano- and micro-scale topographic cues for engineered vascular constructs by electrospinning and imprinted micro-patterns
1
2014
... Alignment, one of the geometric arrangements, also exerts a great effect on cell behaviours. For cell adhesion, it was found that FAK phosphorylation was significantly increased in MSCs on aligned nanofibres compared with random nanofibres.105 For cell spreading, it was found that the cytoskeleton of human umbilical vein ECs on aligned fibres was stretched along the longitudinal direction of the fibre, while it extended in all directions on non-aligned fibres.107 Interestingly, cell alignment showed a more significant effect on micro-aligned scaffolds than on nano-aligned scaffolds.106 For cell migration, compared with random fibres, human adipose-derived stem cells cultured on aligned nanofibres exhibited a faster migration rate and higher synthetic ability of critical ECM molecules, such as collagen type I, tropoelastin, and matrix metalloproteinase-1.108 The effect of the alignment of electrospun polycaprolactone-gelatine ultrafine fibres on cell migration was also studied, and showed that the cell migration rate was faster in the direction of aligned fibres compared with non-aligned fibres.109 In addition, alignment affected cell proliferation and differentiation and was cell-type dependent. For example, aligned scaffold enhanced cell proliferation of keratocytes, while randomly-oriented scaffold enhanced the proliferation of corneal epithelial cells.110 The osteogenic differentiation of MSCs could also be modulated by fibre alignment. Alkaline phosphatase and calcium nodules were evenly distributed along the direction of aligned fibres, while gene expression of osteocalcin and osteopontin was significantly enhanced, indicating that aligned fibres were superior to non-aligned fibres in inducing osteogenic differentiation.107 ...
Effect of fiber alignment in electrospun scaffolds on keratocytes and corneal epithelial cells behavior
2
2012
... Alignment, one of the geometric arrangements, also exerts a great effect on cell behaviours. For cell adhesion, it was found that FAK phosphorylation was significantly increased in MSCs on aligned nanofibres compared with random nanofibres.105 For cell spreading, it was found that the cytoskeleton of human umbilical vein ECs on aligned fibres was stretched along the longitudinal direction of the fibre, while it extended in all directions on non-aligned fibres.107 Interestingly, cell alignment showed a more significant effect on micro-aligned scaffolds than on nano-aligned scaffolds.106 For cell migration, compared with random fibres, human adipose-derived stem cells cultured on aligned nanofibres exhibited a faster migration rate and higher synthetic ability of critical ECM molecules, such as collagen type I, tropoelastin, and matrix metalloproteinase-1.108 The effect of the alignment of electrospun polycaprolactone-gelatine ultrafine fibres on cell migration was also studied, and showed that the cell migration rate was faster in the direction of aligned fibres compared with non-aligned fibres.109 In addition, alignment affected cell proliferation and differentiation and was cell-type dependent. For example, aligned scaffold enhanced cell proliferation of keratocytes, while randomly-oriented scaffold enhanced the proliferation of corneal epithelial cells.110 The osteogenic differentiation of MSCs could also be modulated by fibre alignment. Alkaline phosphatase and calcium nodules were evenly distributed along the direction of aligned fibres, while gene expression of osteocalcin and osteopontin was significantly enhanced, indicating that aligned fibres were superior to non-aligned fibres in inducing osteogenic differentiation.107 ...
... Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
Exploring and engineering the cell surface interface
1
2005
... Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
Harnessing nanotopography and integrin-matrix interactions to influence stem cell fate
1
2014
... Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
Surface roughness and substrate stiffness synergize to drive cellular mechanoresponse
2
2020
... Many remarkable articles have discussed cellular responses to substrate topography.105, 111, 112 In recent years, cell behaviour cross-talk between substrate topography and stiffness has attracted great attention.10 For example, in vascular tissue engineering, MSCs cultured on a micro-grooved, stiff substrate exhibited maximum cell stiffness and smooth muscle-related gene expression of α-actin and h1-calponin while MSCs showed minimum values on a smooth, soft substrate.110 Our group studied the effect of substrate stiffness and topography on the differentiation of AF-derived stem cells, and found that they both regulated the differentiation of AFSCs through a YAP-dependent mechano-transduction mechanism.102 Recently, Hou et al.113 developed large-scale surface roughness gradient hydrogels by soft lithography technology, integrating a broad range of surface roughnesses (200 nm to 1.2 μm for root-mean-squared surface roughness) with tuneable stiffness. They found that MSCs responded to surface roughness in a stiffness-dependent manner by reorganizing the surface hierarchy.113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
... 113 Interestingly, on the soft hydrogel (3.8 kPa) with high surface roughness, both the cellular mechano-response and osteogenesis increased significantly, better than that on the stiff substrates with smooth surfaces. ...
Incorporation of F-MWCNTs into electrospun nanofibers regulates osteogenesis through stiffness and nanotopography
2
2020
... Numerous studies have indicated that increased surface nano-roughness or stiffness of electrospun fibrous scaffolds is conducive for an osteogenic response. Recently, Jahanmard et al.114 incorporated multiwall carbon nanotubes into electrospun polycaprolactone nanofibres to adjust surface nanoroughness and stiffness independently. They found that osteoblast differentiation was improved by both these crucial parameters, yet through different mechanisms.114 All of these studies help us to understand the mechanical and biological responses of bone progenitor cells, and ultimately guide us to design better scaffolds for bone tissue engineering. ...
... 114 All of these studies help us to understand the mechanical and biological responses of bone progenitor cells, and ultimately guide us to design better scaffolds for bone tissue engineering. ...
Cell migration: implications for repair and regeneration in joint disease
1
2019
... Fibrous scaffolds are also commonly being explored to restore the function and organization of dense connective tissues such as tendon, knee meniscus, and cartilage.115, 116 However, the lower porosity of electrospun fibres results in poor cell invasion, hindering cell migration into scaffolds. To test the impact of stiffness of electrospun fibres on cell invasion into a fibrous scaffold, Song et al.117 fabricated stiff and soft electrospun fibrous networks using crosslinked methacrylated hyaluronic acid. Enhanced meniscal fibrochondrocyte invasion and greater collagen deposition were observed on stiffer fibrous networks when compared with softer fibrous networks because of their easy deformation and increase in density under the influence of cell traction. All these results demonstrated that the stiffness of fibrous scaffolds is a key parameter in the design of scaffolds for tissue regeneration. ...
Electrospun nanofibers for regenerative medicine
1
2012
... Fibrous scaffolds are also commonly being explored to restore the function and organization of dense connective tissues such as tendon, knee meniscus, and cartilage.115, 116 However, the lower porosity of electrospun fibres results in poor cell invasion, hindering cell migration into scaffolds. To test the impact of stiffness of electrospun fibres on cell invasion into a fibrous scaffold, Song et al.117 fabricated stiff and soft electrospun fibrous networks using crosslinked methacrylated hyaluronic acid. Enhanced meniscal fibrochondrocyte invasion and greater collagen deposition were observed on stiffer fibrous networks when compared with softer fibrous networks because of their easy deformation and increase in density under the influence of cell traction. All these results demonstrated that the stiffness of fibrous scaffolds is a key parameter in the design of scaffolds for tissue regeneration. ...
Influence of fiber stiffness on meniscal cell migration into dense fibrous networks
1
2020
... Fibrous scaffolds are also commonly being explored to restore the function and organization of dense connective tissues such as tendon, knee meniscus, and cartilage.115, 116 However, the lower porosity of electrospun fibres results in poor cell invasion, hindering cell migration into scaffolds. To test the impact of stiffness of electrospun fibres on cell invasion into a fibrous scaffold, Song et al.117 fabricated stiff and soft electrospun fibrous networks using crosslinked methacrylated hyaluronic acid. Enhanced meniscal fibrochondrocyte invasion and greater collagen deposition were observed on stiffer fibrous networks when compared with softer fibrous networks because of their easy deformation and increase in density under the influence of cell traction. All these results demonstrated that the stiffness of fibrous scaffolds is a key parameter in the design of scaffolds for tissue regeneration. ...
Engineered fibrous networks to investigate the influence of fiber mechanics on myofibroblast differentiation
1
2019
... In addition, the stiffness of fibrous scaffolds also plays an important role in disease development processes or tissue repair processes. For example, our group recently reported that fibre stiffness of polyurethanes and fibre diameters showed a synergistic effect on the differentiation of AFSCs. AFSCs had a strong tendency to differentiate into various regional AF-like cells when cultured on a scaffold with physical cues similar to native AF tissue.102 Davidson et al.118 seeded hepatic stellate cells onto soft or stiff fibrous hyaluronic acid hydrogels, and found that cell spreading, α-smooth muscle actin expression, and formation of multicellular clusters decreased on stiff fibres compared with those on soft fibres, indicating the important influence of fibre stiffness on cell-material interactions and implying its relevance to disease and tissue repair processes. ...
Tissue force programs cell fate and tumor aggression
1
2017
... Accumulating evidence suggests that external mechanical stimulation regulates cellular processes and strongly influences cell fate.119 Stretching, compression, hydrostatic pressure and fluid shear force can affect cell self-renewal, differentiation, senescence and apoptosis, and play an important role in physiological and pathological processes.11 Indeed, cells in living tissues are constantly exposed to various chemical and physical signals, and then integrate these signals to guide cell differentiation specifics, support morphogenetic processes and define tissue architecture during embryonic development.120 Intracardiac fluid forces change cytoskeletal structure and gene expression profiles of cardiac ECs to promote the morphogenesis of cardiac tissues.121 Tensile forces, generated by the stretching of bronchial epithelium during intrauterine respiration, support the development of pulmonary smooth muscle.122 Long-bone morphogenesis is dependent on both local morphogen gradients and mechanical forces which guide cell assembly and promote growth to higher-order structures.123 ...
Biomaterials and engineered microenvironments to control YAP/TAZ-dependent cell behaviour
1
2018
... Accumulating evidence suggests that external mechanical stimulation regulates cellular processes and strongly influences cell fate.119 Stretching, compression, hydrostatic pressure and fluid shear force can affect cell self-renewal, differentiation, senescence and apoptosis, and play an important role in physiological and pathological processes.11 Indeed, cells in living tissues are constantly exposed to various chemical and physical signals, and then integrate these signals to guide cell differentiation specifics, support morphogenetic processes and define tissue architecture during embryonic development.120 Intracardiac fluid forces change cytoskeletal structure and gene expression profiles of cardiac ECs to promote the morphogenesis of cardiac tissues.121 Tensile forces, generated by the stretching of bronchial epithelium during intrauterine respiration, support the development of pulmonary smooth muscle.122 Long-bone morphogenesis is dependent on both local morphogen gradients and mechanical forces which guide cell assembly and promote growth to higher-order structures.123 ...
Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis
1
2003
... Accumulating evidence suggests that external mechanical stimulation regulates cellular processes and strongly influences cell fate.119 Stretching, compression, hydrostatic pressure and fluid shear force can affect cell self-renewal, differentiation, senescence and apoptosis, and play an important role in physiological and pathological processes.11 Indeed, cells in living tissues are constantly exposed to various chemical and physical signals, and then integrate these signals to guide cell differentiation specifics, support morphogenetic processes and define tissue architecture during embryonic development.120 Intracardiac fluid forces change cytoskeletal structure and gene expression profiles of cardiac ECs to promote the morphogenesis of cardiac tissues.121 Tensile forces, generated by the stretching of bronchial epithelium during intrauterine respiration, support the development of pulmonary smooth muscle.122 Long-bone morphogenesis is dependent on both local morphogen gradients and mechanical forces which guide cell assembly and promote growth to higher-order structures.123 ...
Stretch-induced alternative splicing of serum response factor promotes bronchial myogenesis and is defective in lung hypoplasia
1
2000
... Accumulating evidence suggests that external mechanical stimulation regulates cellular processes and strongly influences cell fate.119 Stretching, compression, hydrostatic pressure and fluid shear force can affect cell self-renewal, differentiation, senescence and apoptosis, and play an important role in physiological and pathological processes.11 Indeed, cells in living tissues are constantly exposed to various chemical and physical signals, and then integrate these signals to guide cell differentiation specifics, support morphogenetic processes and define tissue architecture during embryonic development.120 Intracardiac fluid forces change cytoskeletal structure and gene expression profiles of cardiac ECs to promote the morphogenesis of cardiac tissues.121 Tensile forces, generated by the stretching of bronchial epithelium during intrauterine respiration, support the development of pulmonary smooth muscle.122 Long-bone morphogenesis is dependent on both local morphogen gradients and mechanical forces which guide cell assembly and promote growth to higher-order structures.123 ...
Vertebrate skeletogenesis
1
2010
... Accumulating evidence suggests that external mechanical stimulation regulates cellular processes and strongly influences cell fate.119 Stretching, compression, hydrostatic pressure and fluid shear force can affect cell self-renewal, differentiation, senescence and apoptosis, and play an important role in physiological and pathological processes.11 Indeed, cells in living tissues are constantly exposed to various chemical and physical signals, and then integrate these signals to guide cell differentiation specifics, support morphogenetic processes and define tissue architecture during embryonic development.120 Intracardiac fluid forces change cytoskeletal structure and gene expression profiles of cardiac ECs to promote the morphogenesis of cardiac tissues.121 Tensile forces, generated by the stretching of bronchial epithelium during intrauterine respiration, support the development of pulmonary smooth muscle.122 Long-bone morphogenesis is dependent on both local morphogen gradients and mechanical forces which guide cell assembly and promote growth to higher-order structures.123 ...
Sensory innervation in porous endplates by Netrin-1 from osteoclasts mediates PGE2-induced spinal hypersensitivity in mice
1
2019
... In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130 ...
A mechano-regulation model for tissue differentiation during fracture healing: analysis of gap size and loading
1
2002
... In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130 ...
Endothelial cell biomechanical responses are dependent on both fluid shear stress and tensile strain
1
2019
... In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130 ...
The effects of mechanical forces on intestinal physiology and pathology
1
2009
... In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130 ...
Biomechanical signals exert sustained attenuation of proinflammatory gene induction in articular chondrocytes
1
2006
... In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130 ...
Cyclic tensile stress exerts a protective effect on intervertebral disc cells
1
2008
... In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130 ...
Dynamic biomechanical strain inhibits IL-1beta-induced inflammation in vocal fold fibroblasts
1
2007
... In addition to guiding embryonic development, external mechanical stimulation plays a central role in the growth, differentiation and regeneration of cells or tissues. Vigorous exercise and muscular activity can enhance bone mass and bone strength. Conversely, it leads to the phenomenon of bone loss in the absence of mechanical loading, such as long-term bed rest or spaceflight.124 Moreover, only under mechanical conditions, i.e., cyclic compression at the fracture site, the process of bone fracture healing can be replicated.125 Hence, mechanical loading is necessary for the homeostasis, repair and remodelling of native bone tissue. Moreover, in the cardiovascular system, mechanical forces induced by physiological blood flow cause the cytoskeleton proteins of ECs to align with the blood flow direction to effectively reduce blood-flow resistance.126 The mucosal cells in the gastrointestinal tract are subjected to pressure and shear stress when they interact with relatively incompressible endoluminal chyme to maintain their functions.127 Many studies have demonstrated that cyclic tensile strain of a low magnitude and low frequency has an anti-inflammatory effect, increasing anabolic gene expression and matrix synthesis in various cell types, including chondrocytes,128 AF cells129 and fibroblasts.130 ...
Disordered mechanical stress and tissue engineering therapies in intervertebral disc degeneration
1
2019
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Cyclic tensile stress of human annulus fibrosus cells induces MAPK activation: involvement in proinflammatory gene expression
1
2016
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Notochordal cell disappearance and modes of apoptotic cell death in a rat tail static compression-induced disc degeneration model
1
2014
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Excessive mechanical loading promotes osteoarthritis through the gremlin-1-NF-κB pathway
1
2019
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Cyclic tensile strain upregulates pro-inflammatory cytokine expression via FAK-MAPK signaling in chondrocytes
1
2018
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Protective effects of cilengitide on inflammation in chondrocytes under excessive mechanical stress
1
2020
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Endothelial cell activation. A central pathophysiological process
1
1998
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Membrane depolarization is the trigger for PI3K/Akt activation and leads to the generation of ROS
1
2012
... In contrast to moderate mechanical loading, excessive or aberrant mechanical loading has a negative regulatory effect on cell responses. As is well known, mechanical overload is one of the major factors leading to intervertebral disc degeneration, which accounts for low back pain and physical disability.131 Disordered mechanical loading directly affects intervertebral disc cell behaviours. Pratsinis et al.132 found that mechanical overload induced the imbalance of anabolism and catabolism, and caused inflammation of AF cells. Using a rat tail disc degeneration model, Yurube et al.133 showed that prolonged static compression resulted in significant cell loss of nucleus pulposus and increased the expression of apoptosis markers. Articular cartilage degeneration in osteoarthritis has also been attributed to excessive mechanical loading.134 Previous studies have reported that excessive mechanical stimulation increases the expression of pro-inflammatory cytokines and matrix metalloproteinases in chondrocytes, which are related to the inflammatory response and matrix degradation.135, 136 In addition, homeostatic disturbance caused by abnormal shear forces can change the phenotype of ECs from anti-thrombotic to pro-thrombotic, and thus promote platelet adhesion.137 Chatterjee et al.138 reported that pathological shear stress in ECs reduces nitric oxide production and increases low density lipoprotein oxidation and apoptosis, ultimately resulting in vascular inflammation. ...
Integrins: versatility, modulation, and signaling in cell adhesion
2
1992
... A comprehensive understanding of the cellular response to ECM properties is critical for developing suitable biomaterials to mimic natural tissues and increase seeded cell growth. How substrate mechanical signals are translated into intracellular bio-signals needs to be thoroughly elucidated. Cells perceive the mechanical environment through cell-cell adhesion and cell-ECM interactions, mechanosensitive ion channels and their primary cilia.14 Several mechanotransduction pathways have been proven to transduce mechanical cues into biochemical signals including FAK activated by integrins, Rho/RHO-related protein kinase 1 (ROCK) and mitogen-activated protein kinase (MAPK) as mechanoresponsive signalling, and downstream mechanotransduction signalling such as YAP/PDZ-binding motif (TAZ) (Figure 6), as summarized in Hynes’s review.139 ...
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Integrin signaling through FAK in the regulation of mammary stem cells and breast cancer
1
2010
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
The dual kinase complex FAK-Src as a promising therapeutic target in cancer
1
2010
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Matrix stiffness regulates the differentiation of tendon-derived stem cells through FAK-ERK1/2 activation
1
2018
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Biomechanical force in blood development: extrinsic physical cues drive pro-hematopoietic signaling
1
2013
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
The use of nanoscale topography to modulate the dynamics of adhesion formation in primary osteoblasts and ERK/MAPK signalling in STRO-1+ enriched skeletal stem cells
1
2009
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Nanotopography modulates mechanotransduction of stem cells and induces differentiation through focal adhesion kinase
3
2013
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
... Other mechanical cues, including matrix topography, fibre stiffness, and external mechanical stimulation, also affect the gene expression and fate of cells through the above signalling pathways and their interplays. For example, with the increase of FAK phosphorylation, the neuronal differentiation of hMSCs was promoted on equally-spaced nanogratings with widths of 250145 to 500 nm,154 suggesting that FAK phosphorylation is a signal transducer between integrins and the cytoskeleton, transmitting nanotopographical stimuli to the nucleus through intracellular contraction.145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158 ...
... 145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158 ...
ECM compliance regulates osteogenesis by influencing MAPK signaling downstream of RhoA and ROCK
1
2009
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Transduction of mechanical and cytoskeletal cues by YAP and TAZ
1
2012
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Control of cellular responses to mechanical cues through YAP/TAZ regulation
1
2019
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Integrin activation and internalization on soft ECM as a mechanism of induction of stem cell differentiation by ECM elasticity
1
2011
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels
1
2010
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Mechanosensitive ion channels push cancer progression
1
2019
... The transmembrane receptor integrin, which clusters the assembly of adaptor molecules, acts as a link between the ECM and the cytoskeleton.139 When linking to ECM ligands, integrin simultaneously initiates the assembly of FAK, vinculin, paxillin, tensin, and talin to form FAs. After FA assembly, many signalling pathways will be activated to regulate cell fate.140, 141 Changes of FA composition, size and strength often result in actin contractility and trigger activation of signal molecules which subsequently initiate cell behaviours. For instance, Liu et al.142 found that increased matrix elasticity enhanced FAK activity to modulate the phosphorylation of the down-stream molecule extracellular signal-regulated kinase 1/2, promoting tenogenic differentiation of tendon-derived stem cells. FAK then phosphorylates and activates mechanically-responsive signal elements such as MAPK and transforming protein RHOA (Ras homolog gene family member A). MAPK conveys information from the ECM into the nucleus and plays a unique role in cell fate.143 Recent evidence indicated that MAPK activity depended on ECM elasticity.144 With respect to RHOA, it can regulate mechanical feedback by activating ROCK. The RHO/ROCK-mediated signalling pathway appears to play a critical role in cell-ECM mechanotransduction, which is also influenced by matrix elasticity.145 A simultaneous study showed that changes in matrix elasticity altered the RHO/ROCK pathway upstream of changes in the MAPK cascade.146 Collectively, these results suggest that activation of the FAK/RHO/ROCK/MAPK signalling pathway via changes of ECM elasticity may play a critical role in cell behaviour. Other than this classical signalling pathway, a variety of novel mechanosensitive proteins are involved in cellular responses to matrix elasticity, including YAP, piezo, and caveolin-1. YAP activity is sustained by the actin cytoskeleton and this pathway is relevant not only to enabling the cellular response to ECM elasticity,147 but also in mediating cellular responses to external stimuli.148 Caveolin-1 is a structural protein of caveolae which is involved in the activation of integrin during extracellular mechanotransduction. In one study, ECM elasticity affected integrin activity and trafficking to regulate the endocytosis of caveolin-1, which, in turn, influenced cell fate and function.149 Piezo ion channels, which fall into two main categories: piezo1 and piezo2, can be detected in many mechanosensitive tissues.150 Piezo channels are known to sense matrix elasticity and directly lead to the increase of intracellular Ca2+. Then the Ca2+ induces YAP nuclear translocation and transfers extracellular mechanical stimuli into the nucleus.151 Together, these studies demonstrated that cells reorganize their actin cytoskeleton via activation of integrin, FAs and other relevant mechanosensitive molecules to respond to ECM elasticity. ...
Dimensionality and spreading influence MSC YAP/TAZ signaling in hydrogel environments
1
2016
... The methods used to explore the molecular mechanism of how cells sense ECM viscoelasticity are similar to those used in studying how cells sense ECM elasticity. RHO family GTPases are significant regulators of the actin cytoskeleton and of mechanotransduction. It has been found that RHO-mediated contractility and independent actomyosin contractility regulates osteogenic differentiation.68, 152 Increased Rac1 activity was also reported to enhance smooth muscle cell differentiation.153 YAP/TAZ has been considered as key mediators in cell responses to substrate stiffness, such as that in viscoelastic regulation of cell behaviours. For example, stress relaxation of a substrate has been found to lead to increased nuclear translocation of YAP, resulting in increased U2OS cell proliferation.74 Further, the cell spreading, proliferation, and osteogenic differentiation of MSCs was enhanced on gels with faster stress relaxation, which was proven to be mediated through integrin-based adhesion, actomyosin contractility, and nuclear localization of YAP.68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume. ...
The effect of time-dependent deformation of viscoelastic hydrogels on myogenic induction and Rac1 activity in mesenchymal stem cells
1
2014
... The methods used to explore the molecular mechanism of how cells sense ECM viscoelasticity are similar to those used in studying how cells sense ECM elasticity. RHO family GTPases are significant regulators of the actin cytoskeleton and of mechanotransduction. It has been found that RHO-mediated contractility and independent actomyosin contractility regulates osteogenic differentiation.68, 152 Increased Rac1 activity was also reported to enhance smooth muscle cell differentiation.153 YAP/TAZ has been considered as key mediators in cell responses to substrate stiffness, such as that in viscoelastic regulation of cell behaviours. For example, stress relaxation of a substrate has been found to lead to increased nuclear translocation of YAP, resulting in increased U2OS cell proliferation.74 Further, the cell spreading, proliferation, and osteogenic differentiation of MSCs was enhanced on gels with faster stress relaxation, which was proven to be mediated through integrin-based adhesion, actomyosin contractility, and nuclear localization of YAP.68 However, some studies presented different phenomena. On viscoelastic boronate ester hydrogels, the nuclear-to-cytosolic ratio of YAP/TAZ was obviously decreased in NIH-3T3 cells.75 The difference may be due to the difference of viscoelastic gel systems. The former undergoes plastic deformation while the latter exhibits elastic recovery of creep strain and its original shape is restored after elimination of shear stress. In addition, when MSCs were encapsulated in a 3D relaxing boronate-based hydrogel,83 the subcellular localization of YAP/TAZ was different from that on 2D relaxing boronate-based hydrogels. On day 1, YAP/TAZ was mainly located in the cytoplasm, and the ratio of nuclear to cytosolic intensity on YAP/TAZ increased significantly on day 7, consistent with the enhanced cell volume and nuclear volume. ...
Nanotopography-induced changes in focal adhesions, cytoskeletal organization, and mechanical properties of human mesenchymal stem cells
1
2010
... Other mechanical cues, including matrix topography, fibre stiffness, and external mechanical stimulation, also affect the gene expression and fate of cells through the above signalling pathways and their interplays. For example, with the increase of FAK phosphorylation, the neuronal differentiation of hMSCs was promoted on equally-spaced nanogratings with widths of 250145 to 500 nm,154 suggesting that FAK phosphorylation is a signal transducer between integrins and the cytoskeleton, transmitting nanotopographical stimuli to the nucleus through intracellular contraction.145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158 ...
YAP-dependent mechanotransduction is required for proliferation and migration on native-like substrate topography
1
2017
... Other mechanical cues, including matrix topography, fibre stiffness, and external mechanical stimulation, also affect the gene expression and fate of cells through the above signalling pathways and their interplays. For example, with the increase of FAK phosphorylation, the neuronal differentiation of hMSCs was promoted on equally-spaced nanogratings with widths of 250145 to 500 nm,154 suggesting that FAK phosphorylation is a signal transducer between integrins and the cytoskeleton, transmitting nanotopographical stimuli to the nucleus through intracellular contraction.145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158 ...
Cyclic stretching of soft substrates induces spreading and growth
1
2015
... Other mechanical cues, including matrix topography, fibre stiffness, and external mechanical stimulation, also affect the gene expression and fate of cells through the above signalling pathways and their interplays. For example, with the increase of FAK phosphorylation, the neuronal differentiation of hMSCs was promoted on equally-spaced nanogratings with widths of 250145 to 500 nm,154 suggesting that FAK phosphorylation is a signal transducer between integrins and the cytoskeleton, transmitting nanotopographical stimuli to the nucleus through intracellular contraction.145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158 ...
Integrin-YAP/TAZ-JNK cascade mediates atheroprotective effect of unidirectional shear flow
1
2016
... Other mechanical cues, including matrix topography, fibre stiffness, and external mechanical stimulation, also affect the gene expression and fate of cells through the above signalling pathways and their interplays. For example, with the increase of FAK phosphorylation, the neuronal differentiation of hMSCs was promoted on equally-spaced nanogratings with widths of 250145 to 500 nm,154 suggesting that FAK phosphorylation is a signal transducer between integrins and the cytoskeleton, transmitting nanotopographical stimuli to the nucleus through intracellular contraction.145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158 ...
Stretch-activated ion channel Piezo1 directs lineage choice in human neural stem cells
1
2014
... Other mechanical cues, including matrix topography, fibre stiffness, and external mechanical stimulation, also affect the gene expression and fate of cells through the above signalling pathways and their interplays. For example, with the increase of FAK phosphorylation, the neuronal differentiation of hMSCs was promoted on equally-spaced nanogratings with widths of 250145 to 500 nm,154 suggesting that FAK phosphorylation is a signal transducer between integrins and the cytoskeleton, transmitting nanotopographical stimuli to the nucleus through intracellular contraction.145 On native-like substrate topography, YAP was identified as a required signalling factor which influenced cell proliferation and migration.155 Meanwhile in regulation of cell behaviour by external mechanical stimulation, diverse molecular mechanisms were synergistically involved in this complex and precise process. YAP activated by cyclic stretching promotes cell spread and growth,156 while disturbed flow leads to YAP activation which mediates atherogenesis.157 In human neural stem cells, the stretch-activated ion channel piezo1 guides lineage selection.158 ...
Polyelectrolyte multilayers of poly (l-lysine) and hyaluronic acid on nanostructured surfaces affect stem cell response
1
2019
... The effects of topography and viscoelasticity on the fate of human adipose-derived stem cells were studied by combining nanostructures with viscoelastic elements. The results showed that in multi-layer modified nanostructures, the differentiation of human adipose-derived stem cells towards chondrogenesis and osteogenesis was better than that towards adipogenesis.159 By analysing the cell arrangement and neural differentiation of human umbilical cord mesenchymal stem cells and the growth of nerve processes in vitro and in vivo, the effects of elasticity and aligned topography as combined mechanical cues on cell behaviours were studied.160 After 2 weeks of culture in medium lacking a soluble neurogenic inducer, low elasticity and arranged topography of a layered fibrin hydrogel showed a synergistic effect in promoting the differentiation of human umbilical cord mesenchymal stem cells into nerve cells when compared with random fibrin hydrogel or a culture plate. In order to simulate the dynamic extracellular environment of bone marrow cells in vivo, a bone marrow stem cell substitute matrix based on a physically cross-linked hydrogel was designed. Elasticity was regulated dynamically by chemical stimulation. Under frequent mechanical stress, hMSCs on this hydrogel maintained the expression of Stro-1 (a potential marker of pluripotency of MSCs) for more than 20 days, regardless of the elasticity of the matrix.161 Stiffness and surface topography of the substrate are two key characteristics at the cell-substrate interface that affect cell behaviour as mentioned before.10 From all these studies, we can conclude that multi-mechanical factors play a synergistic role in guiding cell behaviour, or that some mechanical factors would play a leading role. ...
Co-effects of matrix low elasticity and aligned topography on stem cell neurogenic differentiation and rapid neurite outgrowth
1
2016
... The effects of topography and viscoelasticity on the fate of human adipose-derived stem cells were studied by combining nanostructures with viscoelastic elements. The results showed that in multi-layer modified nanostructures, the differentiation of human adipose-derived stem cells towards chondrogenesis and osteogenesis was better than that towards adipogenesis.159 By analysing the cell arrangement and neural differentiation of human umbilical cord mesenchymal stem cells and the growth of nerve processes in vitro and in vivo, the effects of elasticity and aligned topography as combined mechanical cues on cell behaviours were studied.160 After 2 weeks of culture in medium lacking a soluble neurogenic inducer, low elasticity and arranged topography of a layered fibrin hydrogel showed a synergistic effect in promoting the differentiation of human umbilical cord mesenchymal stem cells into nerve cells when compared with random fibrin hydrogel or a culture plate. In order to simulate the dynamic extracellular environment of bone marrow cells in vivo, a bone marrow stem cell substitute matrix based on a physically cross-linked hydrogel was designed. Elasticity was regulated dynamically by chemical stimulation. Under frequent mechanical stress, hMSCs on this hydrogel maintained the expression of Stro-1 (a potential marker of pluripotency of MSCs) for more than 20 days, regardless of the elasticity of the matrix.161 Stiffness and surface topography of the substrate are two key characteristics at the cell-substrate interface that affect cell behaviour as mentioned before.10 From all these studies, we can conclude that multi-mechanical factors play a synergistic role in guiding cell behaviour, or that some mechanical factors would play a leading role. ...
Frequent mechanical stress suppresses proliferation of mesenchymal stem cells from human bone marrow without loss of multipotency
1
2016
... The effects of topography and viscoelasticity on the fate of human adipose-derived stem cells were studied by combining nanostructures with viscoelastic elements. The results showed that in multi-layer modified nanostructures, the differentiation of human adipose-derived stem cells towards chondrogenesis and osteogenesis was better than that towards adipogenesis.159 By analysing the cell arrangement and neural differentiation of human umbilical cord mesenchymal stem cells and the growth of nerve processes in vitro and in vivo, the effects of elasticity and aligned topography as combined mechanical cues on cell behaviours were studied.160 After 2 weeks of culture in medium lacking a soluble neurogenic inducer, low elasticity and arranged topography of a layered fibrin hydrogel showed a synergistic effect in promoting the differentiation of human umbilical cord mesenchymal stem cells into nerve cells when compared with random fibrin hydrogel or a culture plate. In order to simulate the dynamic extracellular environment of bone marrow cells in vivo, a bone marrow stem cell substitute matrix based on a physically cross-linked hydrogel was designed. Elasticity was regulated dynamically by chemical stimulation. Under frequent mechanical stress, hMSCs on this hydrogel maintained the expression of Stro-1 (a potential marker of pluripotency of MSCs) for more than 20 days, regardless of the elasticity of the matrix.161 Stiffness and surface topography of the substrate are two key characteristics at the cell-substrate interface that affect cell behaviour as mentioned before.10 From all these studies, we can conclude that multi-mechanical factors play a synergistic role in guiding cell behaviour, or that some mechanical factors would play a leading role. ...
The synergetic effect of hydrogel stiffness and growth factor on osteogenic differentiation
1
2014
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
The effect of growth factor environment on fibroblast morphological response to substrate stiffness
1
2013
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
Substrate stiffness together with soluble factors affects chondrocyte mechanoresponses
2
2014
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
... 164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
The cellular response of nerve cells on poly-L-lysine coated PLGA-MWCNTs aligned nanofibers under electrical stimulation
1
2018
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
Differential and interactive effects of substrate topography and chemistry on human mesenchymal stem cell gene expression
2
2018
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
... 167 ...
Photoactivatable hydrogel interfaces for resolving the interplay of chemical, mechanical, and geometrical regulation of collective cell migration
1
2019
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...
Collaborative action of surface chemistry and topography in the regulation of mesenchymal and epithelial markers and the shape of cancer cells
1
2016
... For example, Tan et al.162 found that bone morphogenetic protein-2 significantly inhibits the growth of C2C12 myoblasts on softer substrates while promoting osteogenesis on harder substrates. Grinnell et al.163 found that, in the presence of growth factors that stimulate the overall contraction of cells, cell diffusion will not occur unless adhesion ligand density and substrate stiffness cause the interaction between cell and substrate to be strong enough to resist and overcome the introverted traction force. Chen et al.164 found that transforming growth factor-β1 increased the stiffness and traction of chondrocytes and enhanced their response to stretching, while interleukin-1β had the opposite effect. In addition, the effect of transforming growth factor-β1 on chondrocyte mechanics was stronger on a 90 kPa matrix, while interleukin-1β was stronger on a 1 kPa matrix. Chang et al.165 found that the adhesion, migration and proliferation of ECs were positively correlated with matrix stiffness. Hepatocyte growth factor had a synergistic effect, and the effect of hepatocyte growth factor on a low-stiffness matrix was more obvious.164 Recently, a photoactivated hydrogel interface has been developed to study collective cell migration under standard conditions or under the influence of chemical and mechanical cues. The additive effect of chemical cues and mechanical cues on the inhibition of annular expansion by up-regulating epithelial topography was observed. In addition, the influence of geometric cues becomes more significant with the decrease of chemical cues.166 Some studies confirmed the synergistic effect between surface chemistry and topography in guiding cell behaviours.167, 168 It was found that the effect of the same chemicals on the regulatory proteins was related to surface topography. At the same time, the decrease of cell spreading observed on the super-hydrophilic surface was proven to be related to the decreased expression of the mesenchymal proteins paxillin and vinculin. The synergism between amphoteric ions and nano-porous topography is sufficient to induce the expression of E-cadherin, indicating that a zwitterionic charge group is a powerful regulator of the cell signalling pathway.169 RNA sequence analysis of hMSCs showed that the interaction between chemical treatment and topography was synergistic in all gene ontology functional groups. The analysis of differentially-expressed genes showed that surface topography had a greater effect on gene expression than surface chemistry.167 ...




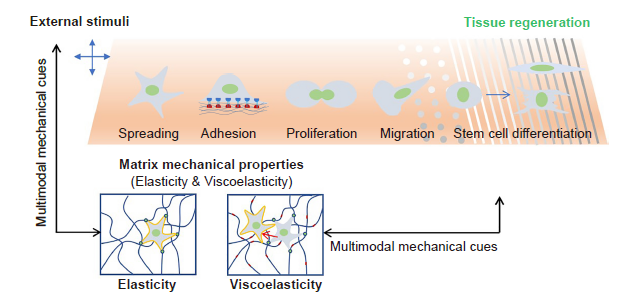
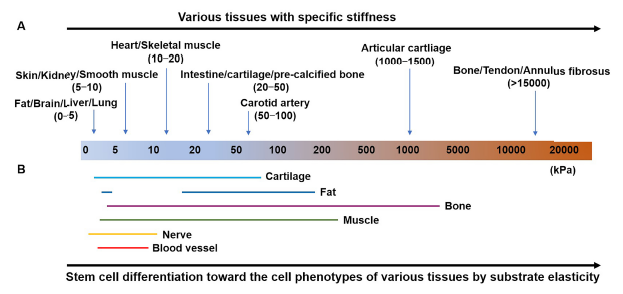
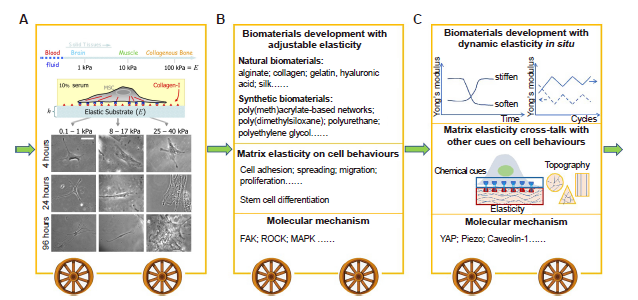
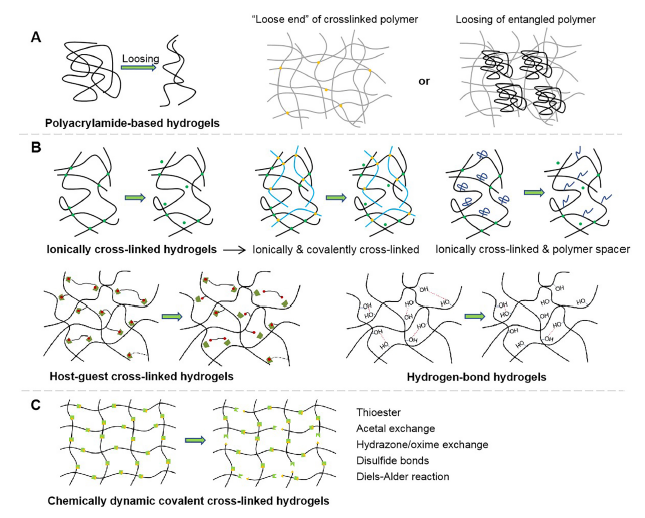
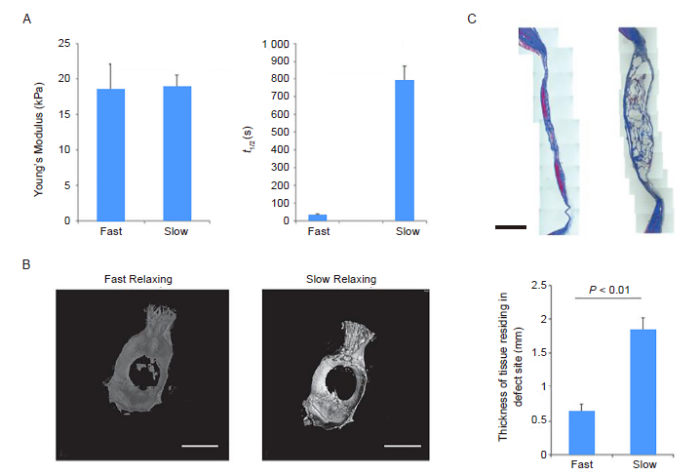
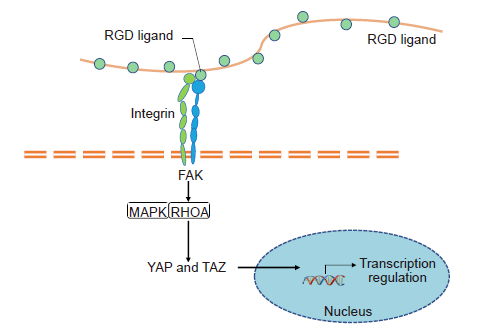

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}